TLRs基因家族在鸭法氏囊中的表达模式及分子进化分析
2018-10-15甘心梦陈凯文刘贺贺李彦莹王继文白丽丽
甘心梦,陈凯文,刘贺贺,李彦莹,王继文,李 亮,白丽丽
(四川农业大学,畜禽遗传资源发掘与创新利用四川省重点实验室,成都 611130)
Toll样受体(toll-like receptors,TLRs)基因家族是机体最早被发现的一类模式识别受体(pattern recognition receptors,PRR),可以准确识别一种或多种病原相关分子模式(pathogen-associated molecular patterns,PAMP),能启动和调节机体的免疫应答,同时也是连接先天性免疫和获得性免疫的关键环节[1-3]。根据脊椎动物TLRs基因家族的胞外结构域可以将其分为TLR1(1、2、6、10)、TLR3、TLR4、TLR5、TLR7(7、8、9)、TLR11、TLR13(13、21)、TLR15八大亚家族[4]。不同物种TLRs基因家族在进化中保持着相似的结构和功能,但其成员组成差别大。果蝇有9种TLRs基因,哺乳动物有13种,其中TLR1~10能在人上特异性表达,小鼠上能表达12种TLRs(TLR1~9、TLR11~13)[5-6],而鸡和鸭均有10种TLRs基因,其中TLR15为禽类特有[7]。禽类TLRs基因家族广泛分布于机体各组织器官中。在鸡不同胚龄组织中,各TLRs基因的表达存在极显著差异(P<0.01)[8]。TLR3和TLR7能介导鸭对病毒感染的先天免疫过程[9]。TLR4主要参与脂多糖(LPS)的识别,注射LPS能刺激鸡小脑的TLR4极显著上调(P<0.01)[10-11]。由此可见,分析组织器官中TLRs家族成员的表达变化可以为探究其功能提供依据。
法氏囊作为禽类特有的中枢免疫器官,是影响其发育、功能形成和免疫功能发挥的大量关键基因的聚集地[12]。在鸡5胚龄时法氏囊开始发育,9~28日龄为生长高峰,56日龄后开始退化[13]。而鸭法氏囊在17周龄时开始退化[14]。研究发现,鸡法氏囊中的TLR3和TLR15、TLR2a和TLR4基因分别在识别马立克氏病毒和新城疫病毒中起着关键作用[15-16]。
基因家族各成员一般来源于祖先基因的基因重复以及分子进化[17-18]。基因重复导致了基因多样化发展,而随着新功能和基因缺失的增加,基因家族出现亚功能化,最终导致进化分歧[19-20]。在其他物种上,FoxO基因家族和Coq1基因家族等都在长期的进化过程中呈现出了不同形式的功能分化[21-22]。为探寻鸭TLRs基因家族各成员的功能差异,本研究拟检测TLRs家族各成员mRNA在鸭法氏囊胚后发育各阶段的相对表达模式,并结合分析启动子区和编码区的系统进化树,以探讨TLRs基因家族表达模式、功能分化与基因分子进化的关系,进一步为TLRs基因家族的研究分析提供基础数据。
1 材料与方法
1.1 试验材料
选取四川农业大学水禽育种场提供的0、2、4和6周龄的健康农华肉鸭(GF2),每周龄公母各3只(共24只),屠宰后采集法氏囊样品,并用液氮研磨后放入-80 ℃超低温冰箱冻存。
1.2 试验方法
1.2.1 鸭法氏囊TLRs的定量引物设计 根据GenBank数据库中鸭TLRsmRNA基因序列,利用Primer Premier 5.0软件设计TLRs基因编码区定量引物(表1),通过PCR扩增、克隆、测序,用Blast程序将测序结果与GenBank数据库中的已知序列进行比对,检测引物的正确性和特异性,包括鸭TLRs基因家族全部10个成员。
1.2.2 鸭法氏囊TLRs的PCR扩增和qRT-PCR反应 采用Trizol法提取鸭法氏囊的总RNA,检测浓度和完整性后,反转录(试剂盒购自TaKaRa公司)合成模板DNA(cDNA),稀释后作为检测模板在荧光定量PCR仪(Bio-Rad)上进行qRT-PCR反应,以β-actin为内参基因,每个样品设3个重复,测定TLRs各成员的相对转录水平。
PCR反应体系为10 μL:模板DNA 1 μL,上下游引物各0.5 μL,PCR Mix 5 μL,ddH2O 3 μL。PCR扩增程序:95 ℃预变性 5 min;95 ℃变性30 s,各基因退火温度如表1所示,退火时间30 s,72 ℃延伸50 s,30个循环;72 ℃最终延伸10 min。
表1鸭TLRs基因和内参基因定量引物信息
Table1InformationofprimersofduckTLRsandreferencegene
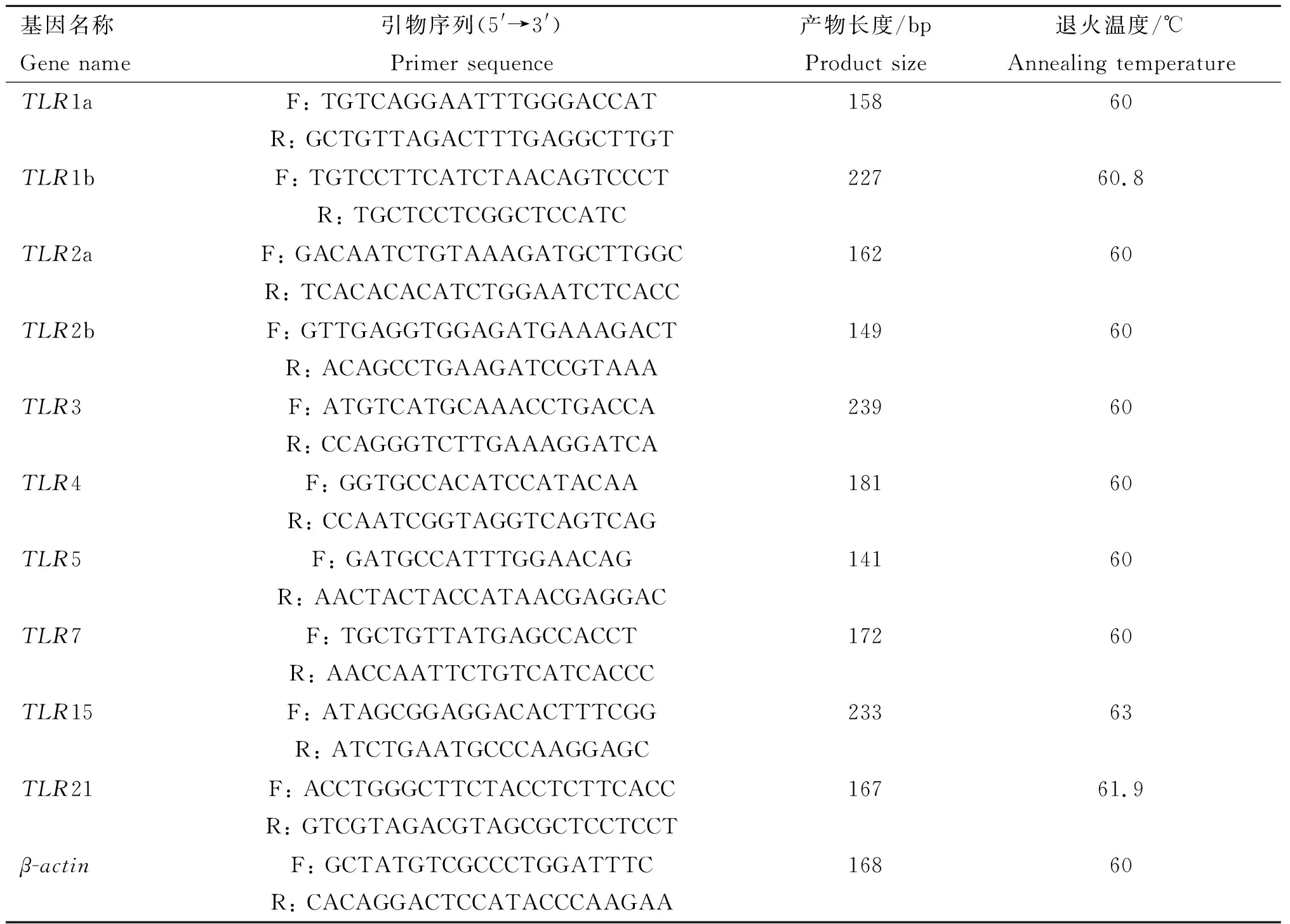
基因名称Gene name引物序列(5'→3')Primer sequence产物长度/bpProduct size退火温度/℃Annealing temperatureTLR1aF: TGTCAGGAATTTGGGACCATR: GCTGTTAGACTTTGAGGCTTGT15860TLR1bF: TGTCCTTCATCTAACAGTCCCTR: TGCTCCTCGGCTCCATC22760.8TLR2aF: GACAATCTGTAAAGATGCTTGGCR: TCACACACATCTGGAATCTCACC16260TLR2bF: GTTGAGGTGGAGATGAAAGACTR: ACAGCCTGAAGATCCGTAAA14960TLR3F: ATGTCATGCAAACCTGACCAR: CCAGGGTCTTGAAAGGATCA23960TLR4F: GGTGCCACATCCATACAAR: CCAATCGGTAGGTCAGTCAG18160TLR5F: GATGCCATTTGGAACAGR: AACTACTACCATAACGAGGAC14160TLR7F: TGCTGTTATGAGCCACCTR: AACCAATTCTGTCATCACCC17260TLR15F: ATAGCGGAGGACACTTTCGGR: ATCTGAATGCCCAAGGAGC23363TLR21F: ACCTGGGCTTCTACCTCTTCACCR: GTCGTAGACGTAGCGCTCCTCCT16761.9β-actinF: GCTATGTCGCCCTGGATTTCR: CACAGGACTCCATACCCAAGAA16860
F.上游引物;R.下游引物
F.Forward primer;R.Reverse primer
qRT-PCR反应体系为20 μL:SYBR@Premix EX TapTM(2×) 10 μL,上下游引物各0.4 μL,ddH2O 7.2 μL,模板DNA 2 μL。反应程序:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火30 s,40个循环;65~95 ℃按0.5 ℃增值进行熔解曲线分析。
1.2.3 构建TLRs编码区和启动子区系统进化树TLRs基因序列均下载自NCBI数据库(http://www.ncbi.nlm.nih.gov/)。使用了人(Homosapiens)、小鼠(Musmusculus)、鸡(Gallusgallus)和鸭(Anasplatyrhynchos)的TLRs编码区序列,选取转录起始位点前2 000 bp作为启动子区序列。利用MEGA 7.0软件的邻接法(Neighbor-Joining),Bootstrap分析重复数为1 000,构建TLRs基因家族系统进化树,其余参数默认不变(表2)。
1.3 统计分析
qRT-PCR结果用Excel 2016整理,采用2-ΔΔC(T)方法计算,结果使用内参基因β-actin进行标准化校正。用Graphpad Prism 7.0绘图,Mev软件进行聚类分析,SPSS 22.0的ANOVA方法进行显著性比较,P<0.05表示差异显著。
2 结 果
2.1 鸭法氏囊胚后发育过程中TLRs的表达
PCR扩增试验结果显示,每个产物均得到单一目的条带,且与预期片段长度一致(图1),说明定量引物特异性较好。通过PCR扩增试验找到每个TLR定量引物的最适熔解温度(表1)。从图2可以看出,TLR2a的表达水平在鸭法氏囊胚后发育的W2和W6时期显著高于W4时期(P<0.05),其余TLRs各成员在鸭法氏囊胚后发育期的表达差异不显著(P>0.05)。
表2系统进化树参考基因信息
Table2Theinformationofreferencegenesofphylogenetictree
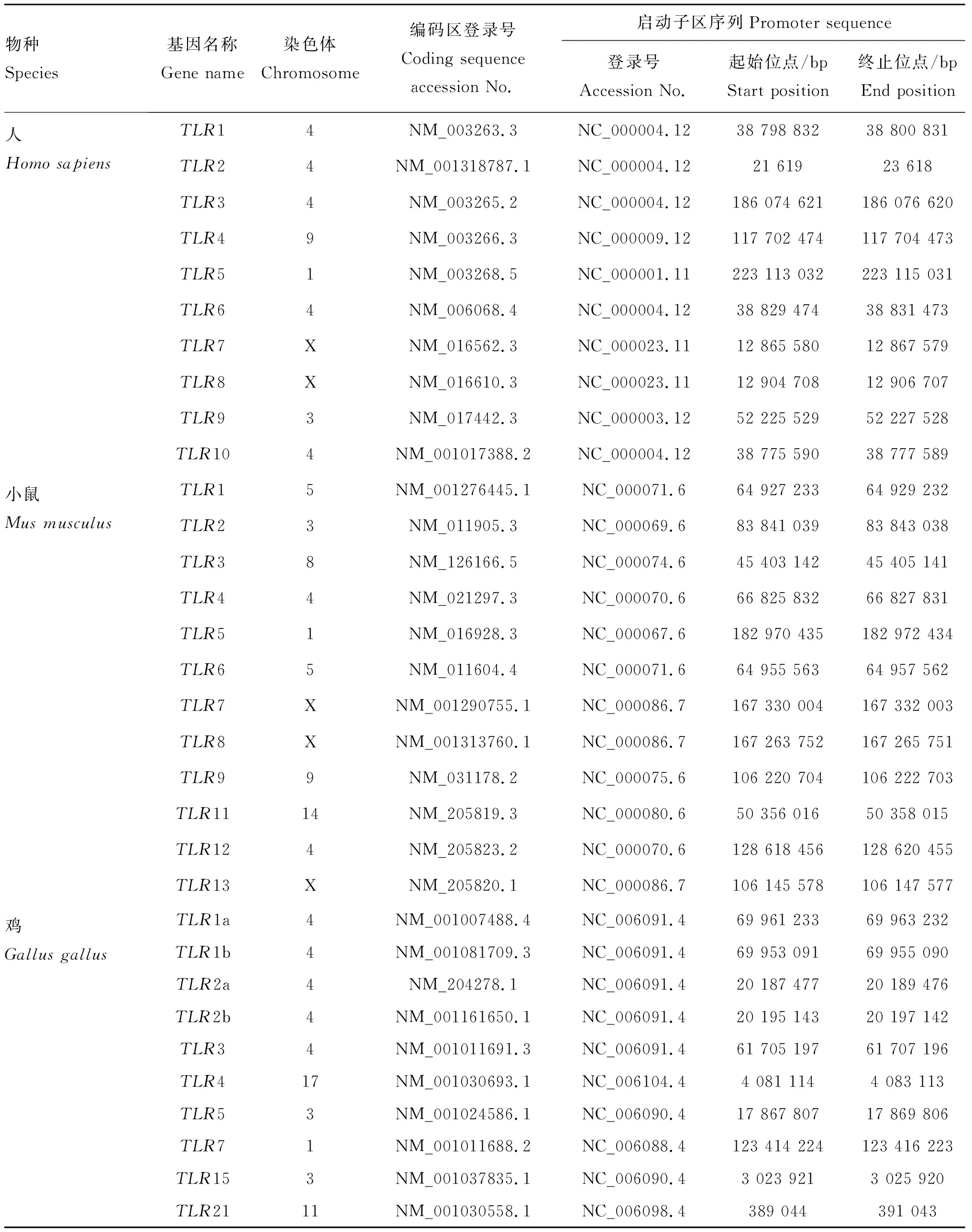
物种Species基因名称Gene name染色体Chromosome编码区登录号Coding sequence accession No.启动子区序列Promoter sequence登录号Accession No.起始位点/bpStart position终止位点/bpEnd position人Homo sapiensTLR14NM_003263.3NC_000004.1238 798 83238 800 831TLR24NM_001318787.1NC_000004.1221 61923 618TLR34NM_003265.2NC_000004.12186 074 621186 076 620TLR49NM_003266.3NC_000009.12117 702 474117 704 473TLR51NM_003268.5NC_000001.11223 113 032223 115 031TLR64NM_006068.4NC_000004.1238 829 47438 831 473TLR7XNM_016562.3NC_000023.1112 865 58012 867 579TLR8XNM_016610.3NC_000023.1112 904 70812 906 707TLR93NM_017442.3NC_000003.1252 225 52952 227 528TLR104NM_001017388.2NC_000004.1238 775 59038 777 589小鼠Mus musculusTLR15NM_001276445.1NC_000071.664 927 23364 929 232TLR23NM_011905.3NC_000069.683 841 03983 843 038TLR38NM_126166.5NC_000074.645 403 14245 405 141TLR44NM_021297.3NC_000070.666 825 83266 827 831TLR51NM_016928.3NC_000067.6182 970 435182 972 434TLR65NM_011604.4NC_000071.664 955 56364 957 562TLR7XNM_001290755.1NC_000086.7167 330 004167 332 003TLR8XNM_001313760.1NC_000086.7167 263 752167 265 751TLR99NM_031178.2NC_000075.6106 220 704106 222 703TLR1114NM_205819.3NC_000080.650 356 01650 358 015TLR124NM_205823.2NC_000070.6128 618 456128 620 455TLR13XNM_205820.1NC_000086.7106 145 578106 147 577鸡Gallus gallusTLR1a4NM_001007488.4NC_006091.469 961 23369 963 232TLR1b4NM_001081709.3NC_006091.469 953 09169 955 090TLR2a4NM_204278.1NC_006091.420 187 47720 189 476TLR2b4NM_001161650.1NC_006091.420 195 14320 197 142TLR34NM_001011691.3NC_006091.461 705 19761 707 196TLR417NM_001030693.1NC_006104.44 081 1144 083 113TLR53NM_001024586.1NC_006090.417 867 80717 869 806TLR71NM_001011688.2NC_006088.4123 414 224123 416 223TLR153NM_001037835.1NC_006090.43 023 9213 025 920TLR2111NM_001030558.1NC_006098.4389 044391 043
(转下页 Carried forward)
1013(scaffold)指TLR21基因在鸭第1013号scaffold上
1013(scaffold) refers to theTLR21 gene on duck’s No.1013 scaffold

图1 TLRs的PCR扩增产物电泳结果Fig.1 The PCR amplification electrophoresis results of TLRs
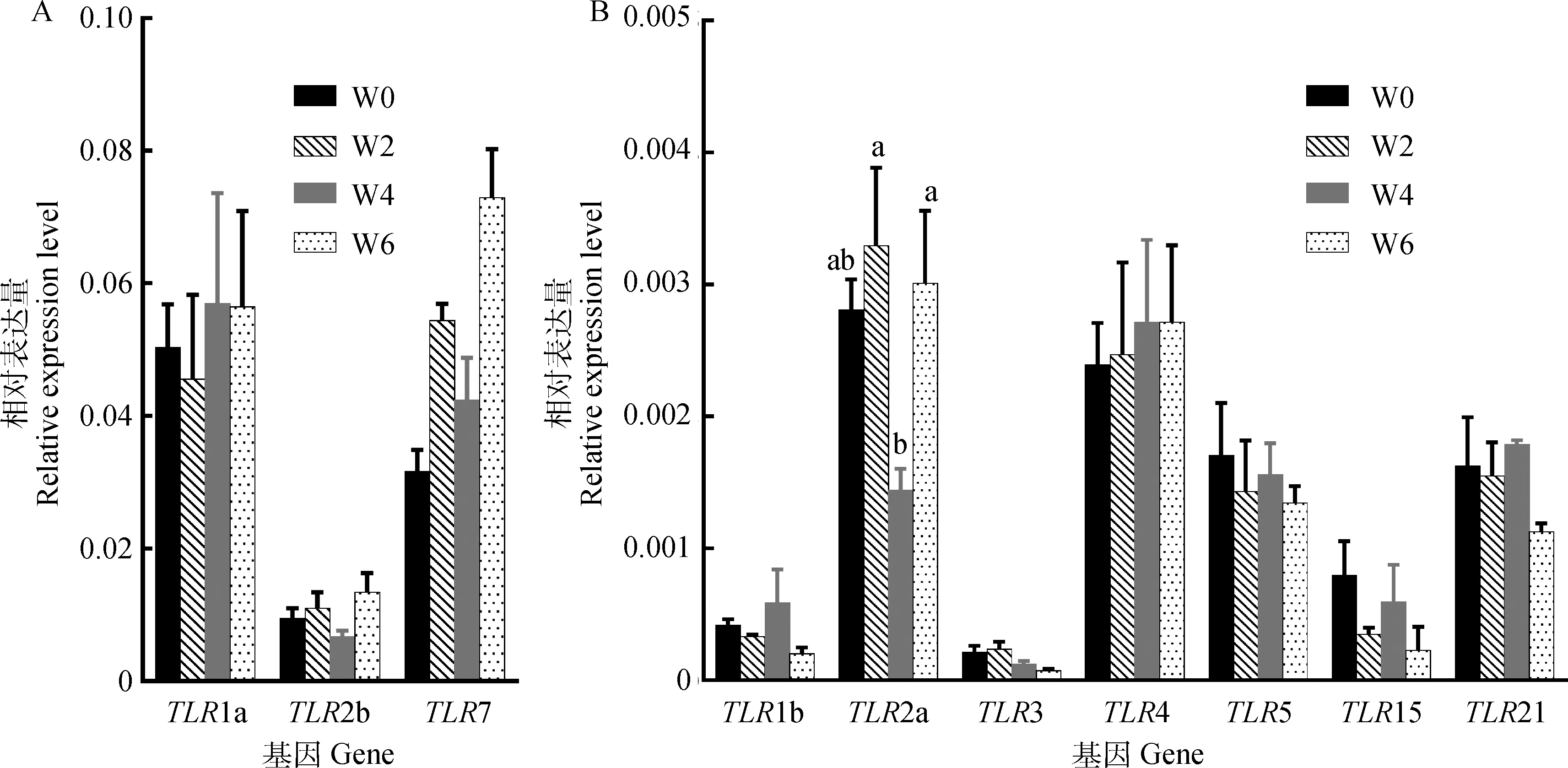
同一基因在4个时期的表达中,不同小写字母表示差异显著(P<0.05)Different lowercase letters indicate significant differences of the same gene expression among 4 periods (P<0.05)图2 鸭法氏囊胚后发育中TLRs的表达情况Fig.2 The expression of TLRs in the postembryonic development stage of duck bursa of Fabricius
2.2 鸭法氏囊TLRs表达聚类分析
如图3,TLR1a、TLR2a和TLR2b的表达模式相近,TLR3、TLR5和TLR15也接近,TLR1b和TLR21的表达模式相近,TLRs基因家族的表达模式可以分为两大类:TLR1a、TLR4、TLR2a、TLR2b和TLR7为一类,其余的TLR3、TLR5、TLR15、TLR1b和TLR21归为一类。

图3 鸭法氏囊胚后发育期TLRs表达量热图Fig.3 The expression heatmap of TLRs in the postembryonic development stage of duck bursa of Fabricius
2.3 TLRs编码区和启动子区的系统进化树
分析TLRs编码区进化树(图4b)可以看出,人、小鼠、鸡和鸭的TLRs基因较为对应的聚在同一分支上。TLR1(1a、1b)和TLR2(2a、2b)聚为一支,与禽类特有的TLR15距离较近;TLR3和TLR5在同一个分支节点上;TLR4和TLR21为单独两分支;鸡和鸭的TLR7与人和小鼠的TLR7、TLR8、TLR9聚在同一支;禽类的TLR21与其他TLRs距离最远。而基于TLRs启动子区构建的系统进化树无明显规律(图4a)。
3 讨 论
在机体免疫应答反应中,TLRs基因家族是极为重要的模式识别受体,通过识别脂多糖、鞭毛蛋白、dsRNA等PAMPs在先天性免疫反应中扮演着重要角色[23]。TLRs基因家族的表达与其行使的功能紧密相关。TLR3和TLR7分别参与了鸡法氏囊对传染性法氏囊病毒和H9N2禽流感病毒的感染应答,表明TLRs在禽类免疫信号感知以及法氏囊免疫功能发挥等方面扮演了重要角色[24-25]。然而,最早发现的果蝇Toll基因被认为是一个发育调控蛋白,鸡TLRs基因在10胚龄到2日龄期间存在明显的表达变化,雏鸭免疫器官中TLRs的表达从33胚龄到1日龄呈显著上升趋势[26-28]。这些研究提示,TLRs基因家族的表达与胚胎发育调控和早期免疫功能有关。在胚胎发育阶段,获得性免疫尚未发育完全,先天性免疫系统已经建立了应对早期致病性攻击的免疫准备[29]。
研究发现,TLRs基因在2~21日龄的雏鸡上并不存在时期表达特异性[27]。在10和30日龄的鸡法氏囊上也并无差异表达基因涉及到TLRs信号通路[30]。本研究结果与现有报道基本一致,除TLR2a外,其余TLRs成员在鸭法氏囊发育阶段的表达均不存在显著差异(P>0.05),提示TLRs基因的表达情况基本稳定,可能不参与鸭法氏囊胚后发育调控过程。另外,禽类TLR2a能识别细菌细胞壁成分,功能与哺乳动物的TLR2和TLR4相近[31]。本研究中,TLR2a在4周龄鸭法氏囊的表达水平显著低于2和6周龄(P<0.05),这可能与该时期鸭法氏囊识别细菌的功能有关,然而仅检测TLRs的表达水平并不能完全代表Toll样受体蛋白的表达情况[32],因此,TLR2a对4周龄鸭法氏囊的发育及功能有何影响还有待进一步研究。
基因家族的进化是由祖先基因通过重复和歧化进化而来,各成员可能具有相同或相关的功能[18]。同时基因的进化也会向着不同物种和种系特异性的方向发展,从而产生特有的基因[33]。Toll基因最初只具有促进机体发育的功能,而由于受到病原体介导的正向选择压力的作用,导致TLRs基因在不同物种的进化中出现差异[19]。本研究中,TLR1a和TLR1b、TLR2a和TLR2b具有较高的同源性,TLR3、TLR5和TLR15的遗传距离较近,TLR21与其余TLRs的遗传关系最远。基因家族中各成员的表达模式比较可以为探究该基因家族的生理生化功能和基因功能分化提供重要信息[34]。如Lopes-Marques等[20]对ACSLs基因家族的研究发现,ACSL1a和1b、ACSL3a和3b、ACSL4a和4b的进化距离较近,其中ACSL3a和3b、ACSL4a和4b功能相似,而ACSL1a和ACSL1b的组织分布表达完全不同,提示其功能存在差异。结合分析TLRs的表达模式与序列进化发现,TLR2a和TLR2b,以及TLR3、TLR5和TLR15具有相似的表达模式和较近的遗传距离,提示其可能具有相似的功能。鸭TLR1a和TLR1b的同源性较高,但表达模式不同,表明可能在基因进化过程中出现了功能分化。
A为TLRs启动子区进化树;B为TLRs编码区进化树,标记为TLRs各成员所在的亚家族A is phylogenetic tree based on TLRs promoter sequence; B is phylogenetic tree based on TLRs coding sequence, the TLRs included in each family are also labeled图4 TLRs系统进化树Fig.4 Phylogenetic trees of TLRs
启动子区位于转录起始位点上游,是调控基因表达、决定基因活动的重要结构,TLRs各成员的表达模式差异是否与启动子区的序列进化有关?为此,本研究对TLRs家族各成员启动子区进行了聚类分析,未发现明显规律,这可能是因为基因的表达与调控是一个复杂过程,涉及到了多个基因和多条信号通路,因此TLRs基因家族表达模式差异可能与启动子区进化无关。
4 结 论
TLRs家族成员在鸭法氏囊中均有表达,且在胚后发育阶段表达变化不明显,提示TLRs家族成员可能不参与鸭法氏囊胚后发育调控过程;TLR2a和TLR2b,以及TLR3、TLR5和TLR15具有相似的表达模式和较近的遗传距离,其功能可能相关;TLR1a和TLR1b虽然具有较高的序列相似性,但表达模式不同,可能出现了功能分化。研究排除了TLRs表达模式与启动子区序列进化的联系。
