废弃铅锌矿石和钨矿砂中可培养细菌多样性分析
2018-10-09邹卫玲冯广达李华平朱红惠
邹卫玲, 冯广达,, 李华平, 朱红惠*
1.广东省微生物研究所, 省部共建华南应用微生物国家重点实验室; 广东省微生物菌种保藏与应用重点实验室; 广东省微生物应用新技术公共实验室, 广州 510070;2.华南农业大学农学院, 广州 510642
金属尾矿中常含有大量的金属元素,长期堆放和风化作用使其中的重金属不断释放,对周围环境造成了严重的污染,也对人们的健康构成了极大的威胁,但重金属污染的治理和修复面临着诸多困难,如效率低、耗时长、二次污染等。研究表明,利用微生物的吸附[1~3]、矿化[4,5]、强化植物富集[6~9]等作用修复重金属污染具有广阔的应用前景[10]。因此,对于这类微生物资源的发掘与保护能够为提高重金属污染的修复效率起到重要的基础支撑作用。目前,相关功能的微生物资源主要分离自受污染的矿区土壤[11~13],而对质地坚硬的废弃矿石及矿砂则关注较少。废弃矿石和矿砂中重金属含量高,营养十分贫瘠,在理化性状上与土壤存在着较大差异,而正是这种极端环境中可能蕴藏着种类丰富的微生物新资源。为此,本研究采用平板稀释涂布法对采自广东省梅州市梅县的废弃铅锌矿石和赣南钨矿砂中的可培养细菌进行分离培养和多样性分析,以期为金属尾矿中微生物新资源的深入挖掘提供参考。
1 材料和方法
1.1 供试材料
废弃铅锌矿石和钨矿砂分别采自广东省梅州市梅县某废弃铅锌矿区和赣南某钨矿区,其理化性质如表1所示。

表1 供试样品的理化性状Table 1 Physicochemical properties of the two samples.
1.2 主要试剂
①R2A培养基[14]:酵母提取物0.5 g,蛋白胨0.5 g,酸水解干酪素0.5 g,葡萄糖0.5 g,可溶性淀粉0.5 g,K2HPO40.3 g,MgSO4·7H2O 0.05 g,丙酮酸钠0.3 g,琼脂粉20 g,双蒸水1.0 L,pH 7.0~7.4;②无磷R2A培养基:去除R2A培养基成分中的K2HPO4,其他成分保持不变;③无磷R2A+Cd2+培养基:向培养基②中添加CdCl2至Cd2+终浓度为2.0 mmol/L。
1.3 分离纯化
分别取10 g粉碎混匀的废弃铅锌矿石和钨矿砂加入到90 mL无菌双蒸水中,140 r/min摇床25℃振荡过夜。用无菌水将菌悬液以10倍梯度逐级稀释,取不同稀释梯度的菌悬液分别涂布于R2A、无磷R2A培养基、无磷R2A+Cd2+培养基上,于30℃培养2周。随后挑取不同形态的单菌落进行进一步纯化培养,并制成甘油浓度为25%(V/V)的甘油菌种,于-80℃冰箱保存备用。
1.4 16S rRNA基因PCR扩增
取适量菌液于12 000 r/min离心3 min后收集菌体,用1 mL无菌双蒸水将菌体涡旋均匀,12 000 r/min离心3 min后弃上清。再加入200 μL的无菌水重悬菌体,涡旋均匀后用液氮冷冻,随后在99℃的固体加热器保持5 min,涡旋30 s,冷冻和加热过程重复2次。最后8 000 r/min离心5 min,取上清液2 μL为模板。采用细菌通用引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-GGTTACCTTGTTACGACTT-3′)进行16S rRNA基因的PCR扩增[15]。反应体系:dNTP(10 mmol/L)0.5 μL,引物(10 μmol/L)各0.5 μL,10×Buffer 2.5 μL,Taq酶(5 U/μL)0.25 μL,模板2 μL,补充ddH2O至25 μL。扩增程序:95℃预变性5 min;94℃变性30 s,55℃退火40 s,72℃延伸1.5 min,共30个循环;72℃延伸15 min。将PCR产物交至上海英骏生物技术公司测序。
1.5 系统进化分析
将菌株的16S rRNA基因序列用生物学软件MAFFT和PHYLIP进行预处理,随后采用软件Mothur以3%的序列差异作为操作分类单元(operational taxonomic unit, OTU)的临界值对序列进行分析[17]。每个OTU中随机选取1株菌的16S rRNA基因序列作为代表上传至GenBank,获取相应的登录号。再在GenBank中选取与之相似度较大的同源模式种序列,采用软件MEGA 5.0以Neighbor-Joining法构建系统进化树,计算模型选择Kimura 2,Bootstrap值设为1 000次。
1.6 生物多样性分析
定义16S rRNA序列同源性大于97%为同一个OTU,采用Mothur计算不同样品中可培养细菌的多样性,包括Simpson多样性指数(D),Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J′)。
计算公式:
Pi表示第i个OTU中的菌株所占比例,S为OTU总数,N为菌株总数,ni表示第i个OTU中的菌株数。
2 结果与分析
2.1 废弃铅锌矿石和钨矿砂中可培养细菌多样性
从废弃铅锌矿石和钨矿砂中共分离细菌152株。其中,98株分离自废弃铅锌矿石,划分为36个OTU,代表序列的GenBank登录号:JQ429448~JQ429483;其余54株分离自钨矿砂,划分为12个OTU,代表序列的GenBank登录号:JQ429484~JQ429495。多样性分析的结果如表2所示,废弃铅锌矿石中可培养细菌的Simpson多样性指数(D)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J′)分别为0.943、3.182和0.888,在多样性及均匀度上均大于钨矿砂,这反映出废弃铅锌矿石中的可培养细菌资源更为丰富。

表2 样品中可培养细菌多样性分析Table 2 Diversity analysis of culturable bacteria isolated from different samples.
2.2 可培养细菌的系统进化分析
废弃铅锌矿石中可培养细菌基于16S rRNA基因序列构建的系统进化树如图1所示,菌株涵盖了5个门(Proteobacteria、Firmicutes、Actinobacteria、Deinococcus-Thermus、Bacteroidetes),以变形菌门(Proteobacteria)为主,约占分离总菌数的74%。Proteobacteria中的Alphaproteobacteria分支分布有15个OTU,占分离菌株总数的34%,主要类群为Sphingomonas和Methylobacterium;Betaproteobacteria分支上分布有5个OTU,占分离菌株总数的28%,Massilia属为主要类群;Gammaproteobacteria中分布有6个OTU,占分离菌株总数的12%,主要类群为Lysobacter和Acinetobacter;Deinococcus-Thermus分支上分布了4个OTU(9PNM-15等),由12株菌组成,约占分离菌株总数的12%,均从属于Deinococcus属;Actinobacteria分支上分布了3个OTU(1PNM-16等),涵盖了5株菌,以Kocuria和Arthrobacter的菌株为主;Firmicutes分支上仅分布了1个OTU,由2株菌组成,为Staphylococcus属的菌株;Bacteroidetes分支上分布了2个OTU,由3株菌组成,其中9NM-4与HymenobacterelongatusVUG-A112T具有较近的进化距离。值得注意的是,9PNM-21与MicroscillamarinaATCC 23134T聚成一簇,但16S rRNA基因序列与已有模式种最大相似度仅为85.5%(表3),可能表示其为Cytophagaceae科内的一个新属。
钨矿砂中可培养细菌基于16S rRNA基因序列构建的系统进化树如图2所示,菌株涵盖了3个门(Proteobacteria、Firmicutes、Actinobacteria),其中变形菌门(Proteobacteria)为优势类群,约占分离菌株总数的81%。Alphaproteobacteria分支上分布有5个OTU,由22株菌组成,占分离菌株总数的37%。Methylobacterium属为其中的优势类群,涵盖了3个OTU(20株菌);其余的2个OTU则分别与Microvirga和Sphingomonas形成稳定的分支。Betaproteobacteria分支上共分布了3个OTU(6NM-7等),包括了21株菌,占分离菌株总数的39%,其中的优势类群为Massilia和Ralstonia。此外,还发现了与Herbaspirillum属亲缘关系较近的菌株;Actinobacteria分支上分布了3个OTU,共9株菌,占分离菌株总数的17%,分别与Streptomyces、Microbacterium和Curtobacterium形成稳定的分支。Firmicutes分支上仅分离到了1个菌株,从属于Bacillus。综上所述,废弃铅锌矿石中的可培养细菌涵盖了5个门、7个分支,以Massilia、Methylobacterium、Deinococcus和Sphingomonas为主要类群。而钨矿砂中的可培养细菌涵盖了3个门、4个分支,以Methylobacterium、Massilia、Ralstonia和Microbacterium为主要类群。Alphaproteobacteria、Betaproteobacteria、Actinobacteria和Firmicutes 4个分支在2种样品中均有分布,而Gammaproteobacteria、Deinococcus-Thermus、Bacteroidetes仅在废弃铅锌矿石中分离获得。2种供试样品中的可培养细菌均以变形菌门(Proteobacteria)为主,占比均在70%以上,而在属水平上分布情况分析发现Methylobacterium、Massilia、Sphingomonas和Herbaspirillum4个属的菌株在2种样品中均有分布,其他属均分布在不同样品中,表明2种金属尾矿样品中的微生物种群在组成上存在较大差异。

图1 基于废弃铅锌矿石中可培养细菌16S rRNA基因序列构建的NJ系统进化树Fig.1 Phylogenetic tree of the bacteria isolated from abandoned lead-zinc ore constructed by the Neighbor-Joining method based on the 16S rRNA gene sequences.注:圆括号中为该OTU代表菌株的16S rRNA基因序列登录号;方括号内为该OTU所代表的菌株数量;分支上的数值代表自举值。
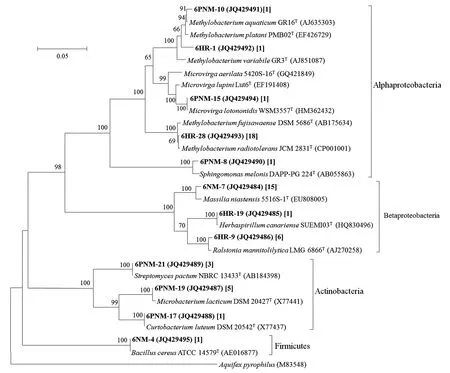
图2 基于钨矿砂中可培养细菌16S rRNA基因序列构建的NJ系统进化树Fig.2 Phylogenetic tree of the bacteria isolated from sands of tungsten mine constructed by the Neighbor-Joining method based on the 16S rRNA gene sequences.注:圆括号中为该OTU代表菌株的16S rRNA基因序列登录号;方括号中为该OTU所代表的菌株数量;分支上的数值代表自举值。
2.3 废弃铅锌矿石和钨矿砂中的疑似新分类单元
根据Kim等[17]的建议将16S rRNA基因序列相似度98.65%作为种间的临界值,在分离的菌株中共发现了疑似新分类单元16个,16株菌与已发表的模式种16S rRNA基因序列最大相似度为85.5%~97.9%。其中,最大相似度为97%~98%、96%~97%的菌株各5株,而最大相似度≤96%的有6株(表3)。仅2个疑似新分类单元(6HR-1和6NM-7)分离自钨矿砂,其余14个均分离自废弃铅锌矿石,废弃铅锌矿石中新分类单元发现率高达14%。目前,采用多相分类学技术手段已完成其中8个新物种的鉴定和命名,即Massiliaputida、Sphingomonasspermidinifaciens、Lysobactermobilis、Acinetobacterguangdongensis、Sphingomonasguangdongensis、Sphingomonasgimensis、Sphingo-monasdifficilis、Deinococcusmetalli[18~25]。

表3 疑似新分类单元的16S rRNA基因序列同源性Table 3 16S rRNA gene sequences homology of the 16 potential new taxa.
注:-表示数据未发表。
2.4 不同分离培养基对可培养细菌类群的影响
不同培养基分离的可培养细菌在门水平上的分布情况如表4所示,R2A培养基分离的菌株涵盖了5个门,无磷酸盐R2A培养基分离的菌株涵盖了4个门,无磷R2A+Cd2+培养基分离的菌株仅涵盖了1个门(Proteobacteria)。从菌株在门水平上的分布可知R2A中磷酸盐的去除对分离的可培养细菌影响较小。而添加2 mmol/L Cd2+使可培养细菌的多样性降低,仅分离到Proteobacteria中3个属的菌株(Methylobacterium、Ralstonia、Herbaspirillum),且均为革兰氏阴性菌,其能够耐受2 mmol/L Cd2+,是金属尾矿中的优势种群。

表4 不同培养基分离获取的细菌在门水平分布情况Table 4 Bacteria distribution in phyla obtained from different media.
3 讨论
废弃铅锌矿石和钨矿砂具有富含重金属(如Cd、Pb、Zn等)、质地坚硬、有机质和水分含量低、营养相对贫乏等极端环境的特点,这种极端环境中可能蕴藏着丰富的具有重金属生物修复潜力的微生物新资源,了解其可培养细菌多样性有助于从中发掘新的资源。研究表明,采用贫营养的培养基有助于从贫瘠环境中分离出种类更多的细菌[26]。R2A培养基中碳源种类丰富,但含量相对较低,属于贫营养型培养基,与金属尾矿营养贫瘠的特征相符,且R2A培养基中含有的丙酮酸钠具有抗氧化作用,能提高微生物的可培养性[27]。Edenborn和Sexstone[28]的研究表明采用R2A培养基分离土壤中的细菌,物种丰富度要显著大于TSA、PIA培养基。因此,本研究采用以R2A为基础的3种培养基研究了废弃铅锌矿石和钨矿砂中可培养细菌多样性。
本研究发现废弃铅锌矿石中可培养细菌的多样性和新分类单元分离率均大于钨矿砂,这可能是由钨矿砂中的Cd含量较高、营养成分相对贫瘠所致。已有的研究也表明重金属Cd能降低可培养细菌对碳源的利用及其生物多样性[29,30]。2种金属尾矿中的可培养细菌均以变形菌门(Proteobacteria)为主,分别占分离菌株总数的74%和81%,根据不同门的革兰氏染色特征可知金属尾矿中的可培养细菌主要为革兰氏阴性菌。本研究还发现仅Methylobacterium、Massilia、Sphingomonas和Herbaspirillum4个属在2种金属尾矿中均有分布,其他属的菌株均分布在不同的样品中,表明这2种材料中的可培养细菌类群组成差异较大,可能与其组成成分及所处环境条件不同有关。与本研究结果相类似,已有的研究也表明Methylobacterium、Massilia、Sphingomonas、Herbaspirillum广泛分布在受污染的土壤[31]和植物体[32,33]中,在维持生态系统稳定方面发挥着重要的作用。
为了进一步了解添加Cd2+对分离的可培养细菌类群的影响,以及供试材料中重金属耐受性较强的细菌类群组成,本研究采用R2A、无磷R2A和无磷R2A+Cd2+培养基对金属尾矿进行了可培养细菌的分离。由于Cd2+与培养基中的磷酸盐易反应产生沉淀,故设置了去除K2HPO4的无磷酸盐R2A培养基作为对照。研究结果表明,K2HPO4的去除对于样品中可培养细菌的分离种类的影响相对较小,可能是由于R2A培养基中的其他营养成分中含有的有机磷源满足了细菌的生长需求。在添加了2 mmol/L Cd2+的无磷培养基中仅分离到Proteobacteria门3个属的菌株,表明重金属Cd2+降低了可培养细菌的多样性。而湛方栋等[34]的研究结果也表明1 mmol/L和2 mmol/L的Cd2+能显著抑制根际土壤微生物的生长。本研究采用的2 mmol/L Cd2+浓度可能超出了2种尾矿样品中大部分可培养细菌的耐受能力,仅Methylobacterium、Herbaspirillum和Ralstonia3个属的菌株对Cd2+表现出较强的耐受性,均为革兰氏阴性菌。与本研究结果相似,付瑾等[35]从江西德兴铜矿土壤中分离到了能够耐受4 mmol/L Cd2+的革兰氏阴性菌Ralstoniapickettii。Zhang等[36]构建了铅锌尾矿定植植物根际耐受0.5 mmol/L Cd2+的可培养细菌16S rRNA基因克隆文库,分析发现其中主要类群也为革兰氏阴性菌。近年来,植物内生菌中也发现了耐受重金属的Methylobacterium、Herbaspirillum、Ralstonia等属的菌株,其在增强植物抗逆性、提高植物富集重金属能力等方面展现了良好的应用效果[37, 38]。
研究表明,传统培养方法能够培养的微生物不足实际数量的0.1%[39],因而,近年来,免培养的高通量测序技术被广泛应用于微生物多样性研究中。本研究未采用该方法主要是受限于废弃铅锌矿石和钨矿砂的总DNA提取存在技术上的困难。尽管高通量测序技术能更加全面的反映样品中微生物的多样性,但传统的分离培养方法在获取功能微生物资源方面仍具有不可替代的作用,其优势在于能够获取目的菌株,并可在此基础上开展相关的生物学特性和功能的研究。也有研究表明传统培养法比免培养的分子生物学技术得到的生物多样性结果能更真实地反映重金属污染程度[40]。因此,在后续的研究中应注重将传统培养方法与免培养的分子生物学技术进行有机结合,既有助于更加全面客观地揭示金属尾矿中的微生物多样性,又能从中获取更多新的功能微生物资源。
