普通小麦 TaTFL1基因的克隆及表达特性研究
2018-10-09何庆梦杨明明李晓燕赵万春
何庆梦,高 翔,2,杨明明,2,董 剑,2,李晓燕,2,赵万春,2
(1.西北农林科技大学农学院,陕西杨凌 712100; 2.陕西省小麦工程技术研究中心/陕西省小麦新品种培育工程研究中心,陕西杨凌 712100)
小麦产量由单位面积穗数、穗粒数和粒重构成。由于受品种遗传特性和坏境条件的制约,粒重的增加幅度相对有限;而穗粒数的变异相对较大,所以通过提高穗粒数来提高小麦单产的潜力大于通过增加粒重的潜力[1]。而穗粒数的形成主要取决于每穗分化小穗数、每穗分化小花数和分化小花的结实数,其中每穗实际分化的小穗数对穗粒数的形成具有重要意义。前人在对控制小穗数基因的研究中发现, Photoperiod-1( Ppd-1)基因能够通过抑制FLOWERINGLOCUST(FT)基因的表达来增加小麦穗粒数[2]。而拟南芥 Terminal Flower 1(TFL1)与FT基因也表现出了明显的相互抑制,在拟南芥和水稻等植物中过表达 TFL1及其同源基因也表现出了成花数的增加[3-4]。由于 Ppd-1与 TFL1基因都能抑制FT基因的表达,因此 TaTFL1也可能对小麦的穗部发育及其小穗数的形成具有重要的作用。
迄今,对 TFL1基因的研究主要是在拟南芥和水稻等模式植物中进行。 TFL1基因在拟南芥中被首次发现,Shannon等[5]通过对拟南芥进行EMS诱变,得到了顶端成花的 tfl-1( terminal flowerl-1)突变体,该突变体提早开花,花序发育受到抑制,且花序被顶花和1~2个侧花所组成的花序结构所终止。TFL1被认为在茎端阻止接受LEAFY(LFY)、APETALA1(AP1)传导的信号,又或是延迟LFY、AP1的上调,以此来抑制LFY、AP1的活性,形成合适的花序结构;LFY、AP1则在花分生组织顶端的外围抑制 TFL1的转录,形成并维持花分生组织的特性[6-8]。在对黑麦草的研究中发现, LpTFL1基因在除成熟的种子外的组织中都有表达,并受到温度与长日照的调控[9]。另外,长日照条件也能诱导 TFL1在植株的营养顶端分生组织中产生较弱水平的表达,随后其表达开始逐渐增强[12]。而水稻中的4个 TFL1同源基因则分别表现出不同的时空表达模式,且有3个基因主要在次级分生组织中表达, Oscen1基因在各时期各组织中都有微弱的表达;而 Oscene2在根和黄化苗中的表达量较强; Oscen3在除叶片外的其他组织表达量较高; Oscen4基因在叶片和叶鞘中未检测到表达,而在根部与黄化苗中表达强烈,在花序和幼苗中表达较弱[11]。Serrano等[3]对拟南芥 TFL1的5′和3′端的顺式调控元件进行研究,发现其表达是基于不同的顺式调控区,且3′端的编码序列对 TFL1的表达最为重要。Guan等[12]在探究激素对拟南芥花序发育影响的研究中发现,细胞分裂素和生长素可能通过诱导分生组织特征基因 TFL1的表达来调节拟南芥花序的形态建成及发育过程,表明激素也参与调控 TFL1基因的表达。这些研究表明, TFL1及其同源基因的表达受诸多因素的影响,在不同植物中存在一定的表达模式差异。
TFL1及其同源基因在植物开花及花序结构的发育中具有重要的作用,且该基因控制植物开花和花序结构的机制可能相对保守[13]。但小麦中尚未见到对该基因的表达特征的相关研究报道。本研究通过克隆小麦第五同源群上 TaTFL1部分同源基因,并利用生物信息学技术对其序列组成、结构特征及进化关系进行初步分析,对启动子区顺式作用元件进行预测,进一步运用qRT-PCR技术分析 TaTFL1在小麦不同组织中的表达特征,以期为进一步探究该基因的功能机制奠定理论基础。
1 材料与方法
1.1 供试材料
普通小麦中国春(CS)由西北农林科技大学小麦品质实验室提供,部分种子在室内培养,在种子萌发后的2、5和10 d分别收集幼根和叶片,并用液氮处理后置于-80 ℃保存备用。同时以室内培养的中国春的幼苗为材料,采用CTAB法提取基因组DNA。部分中国春的种子种植于田间,自小麦种植后分别采集1~2 cm、2~4 cm、4~5 cm和5~6 cm幼穗,同时采集花前穗下茎、颖壳和旗叶以及花后5、10和15 d的穗下茎、颖壳、籽粒和旗叶,用液氮速冻后置于-80 ℃保存备用。
1.2 RNA的提取及cDNA第一链的合成
RNA的提取按照RNAprep Pure Plant Kit(天根,北京)说明书进行,并通过琼脂糖凝胶电泳检测所提取RNA的完整性,使用NanoDropTMOne(Thermo Scientific,美国)测定RNA的浓度。然后根据PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa,日本)说明书合成cDNA第一链。
1.3 TaTFL1基因及其启动子的克隆
根据NCBI(https://www.ncbi.nlm.nih.gov/)已公布的拟南芥 TFL1基因(GenBank登录号:NM_120465.3)序列在Ensembl plants(http://plants.ensembl.org/Triticum_aestivum/Tools/Blast?db=core)进行在线比对,利用比对的最优结果设计特异性引物(表1),以中国春gDNA及cDNA为模板进行扩增。PCR体系与程序参考2×Es Taq MasterMix(康为世纪,北京)说明书,其中循环数为30,退火温度为62 ℃,延伸时间为5 min。PCR产物回收后与克隆载体pEASY-T1(全式金,北京)连接,转化,挑选阳性克隆送公司测序,测序所得结果使用NCBI-BLASTN(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)及DNAMAN 5.0进行序列比对分析。
在Ensembl plants中搜索得到 TaTFL1基因5′端上游2 500 bp大小的序列,并根据其保守区设计引物TP-F和TP-R(表1)进行扩增。PCR体系与程序参考2×Es Taq MasterMix(康为世纪,北京)说明书,其中循环数为30,退火温度为60 ℃,延伸时间为2 min。利用Promoter 2.0 Prediction Server(http://www.cbs.dtu.dk/services/Promoter/)在测序结果中预测启动子区域,再利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对预测的启动子区域进行顺式调控元件分析。
1.4 TaTFL1基因的生物信息学分析
首先将测序结果提交至NCBI-ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/),预测 TaTFL1基因的ORF,利用NCBI-CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/docs/cdd_search.html)预测该基因编码蛋白的结构域。然后,利用GSDS(http://gsds.cbi.pku.edu.cn/)预测基因外显子-内含子结构。进一步,利用ExPASy-ProtParam(https://web.expasy.org/protparam/)、ExPASy-ProtScale(https://web.expasy.org/protscale/)、Cell-PLoc 2.0( http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)、CBS-SignalP(http://www.cbs.dtu.dk/services/SignalP/)和CBS-ProtFun(http://www.cbs.dtu.dk/services/ProtFun/)等分别预测该基因编码蛋白的理化性质、疏水性、亚细胞定位、信号肽及功能;利用NCBI-BLASTP(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=Blast Search&LINK_LOC=blasthome)对所得序列进行同源搜索,获得不同植物中TFL1同源蛋白序列及其登录号,再通过DNAMAN 5.0和MEGA 6.0等软件对TaTFL1同源蛋白进行序列比对及NJ系统进化树的构建,bootstrap值设置为1 000,其他均采用软件默认参数。
1.5 TaTFL1基因表达分析
由于 TaTFL1基因三个部分同源基因序列的一致性很高,因而未能设计出针对每一个基因组进行荧光定量的引物,而是根据三者共同的保守序列设计了共同的荧光定量引物,为避免cDNA中有未除尽的基因组DNA的干扰,荧光定量引物按照跨两个外显子区进行设计,以小麦Actin基因(GenBank登录号:AB181991.1)作为内参基因,引物信息见表1。在ABI 7300 Plus Real Time PCR System(ABI,美国)进行qRT-PCR,体系与程序按照SYBR○RPremix Ex TaqTMII(Tli RNaseH Plus)(TaKaRa,日本)说明书进行,其中循环数为40,退火温度为60 ℃,退火时间为30 s。每个样品设置3次生物学重复及3次技术重复,利用 2-ΔΔCT法[14]计算 TaTFL1基因的相对表达量。
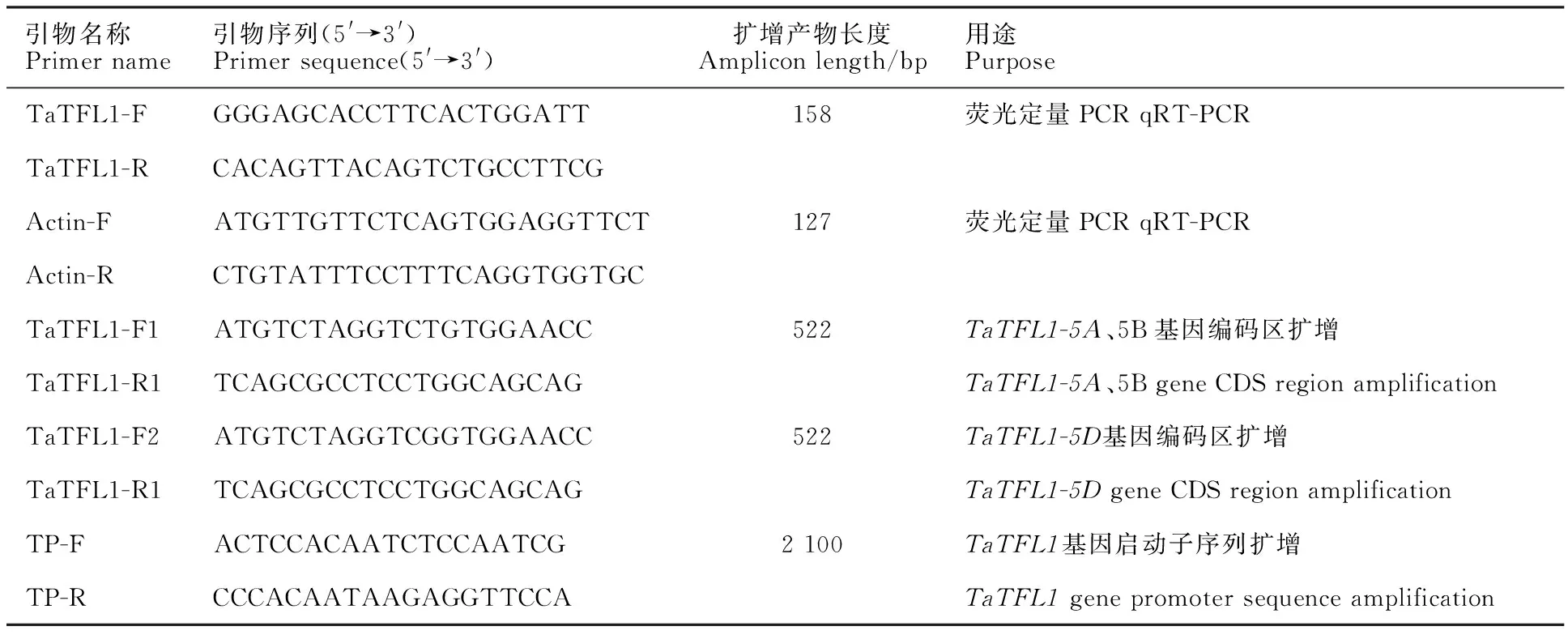
表1 本研究所用的引物信息Table 1 Primers used in this study
2 结果与分析
2.1 中国春 TaTFL1基因的获得及序列特征的初步分析
通过在NCBI中搜索 TFL1基因,获得了拟南芥 TFL1基因的mRNA,将此序列在Ensembl plants中与小麦基因组序列进行比对,获得5AL、5BL和5DL上的 TaTFL1基因。以中国春的gDNA和cDNA为模板,同样以 TaTFL1-F1/ TaTFL1-R1和 TaTFL1-F2/ TaTFL1-R2为引物进行扩增,分别从gDNA和cDNA中扩增得到1 200 bp和520 bp左右的目的条带(图1A)。

M:Trans 2K DNA marker;1: TaTFL1-5A和 TaTFL1-5B cDNA扩增产物; 2: TaTFL1-5D cDNA扩增产物; 3: TaTFL1-5A和 TaTFL1-5B gDNA扩增产物; 4: TaTFL1-5D gDNA扩增产物; 5: TaTFL1启动子扩增产物M:Trans 2K DNA marker; 1:The amplicon of TaTFL1-5A and TaTFL1-5B from cDNA; 2:The amplicon of TaTFL1-5D from cDNA; 3:The amplicon of TaTFL1-5A and TaTFL1-5B from gDNA; 4:The amplicon of TaTFL1-5D from gDNA; 5:The amplicon of TaTFL1 promoter.
将本研究中所获得的3条序列分别命名为 TaTFL1-5A、 TaTFL1-5B和 TaTFL1-5D,并提交至NCBI数据库,得到的序列登录号分别为MF805802.1、MF805803.1和MF805804.1。GSDS预测结果表明, TaTFL1-5A、 TaTFL1-5B和 TaTFL1-5D与其他物种的 TFL1同源基因都具有4个外显子和3个内含子,三者之间只在第一外显子和第二外显子处存在少量的碱基数量差异。将 TaTFL1-5A、 TaTFL1-5B和 TaTFL1-5D的CDS序列及其预测的蛋白质进行相互比对发现彼此之间只存在个别碱基的差异,且大部分碱基差异并未引起氨基酸的改变,除了第190位的碱基差异使与之对应的第64位氨基酸残基在TaTFL1-5D蛋白中表现为M(甲硫氨酸)而TaTFL1-5A与TaTFL1-5B为L(亮氨酸)(图2B)。
2.2 TaTFL1基因编码蛋白的生物信息学分析
利用NCBI-ORF Finder对 TaTFL1基因的ORF进行预测,发现 TaTFL1-5A、 TaTFL1-5B和 TaTFL1-5D均含有一个522 bp的ORF,编码173个氨基酸。使用NCBI-CDD对 TaTFL1基因编码蛋白的保守结构域进行分析,发现TaTFL1-5A、TaTFL1-5B和TaTFL1-5D都具有典型的BEBP家族结构域(图3)。由ExPASy-ProtParam预测结果可知, TaTFL1基因编码蛋白为不稳定蛋白。ExPASy-ProtScale分析结果表明, TaTFL1基因编码蛋白属于兼性蛋白。通过CBS-SignalP对TaTFL1蛋白的信号肽进行预测,结果显示,TaTFL1蛋白不存在信号肽,为非分泌蛋白。Cell-PLoc 2.0的预测结果表明,TaTFL1蛋白定位于细胞质中。通过CBS-Protfun2.2对编码蛋白进行功能预测,结果显示,TaTFL1蛋白在转录、转录调控、免疫应答及生长因子方面的几率均超过0.5,且行使转录调控功能的可能性最大, TaTFL1-5A、TaTFL1-5B和TaTFL1-5D蛋白的几率分别为0.613、0.613和0.705;三者属于酶的可能性均达到了0.46~0.47,几率均大于1.6,表明TaTFL1蛋白可能属于酶。
A:ATG 和 TGA分别代表起始密码子和终止密码子,黑色方框和直线分别代表外显子和内含子,数字表示外显子和内含子的长度(bp)。A:ATG and TGA represent initiation codon and termination codon,respectively. Black box and lines represent exonic and intronic regions,respectively. Numbers indicate the length of exons and introns(bp).

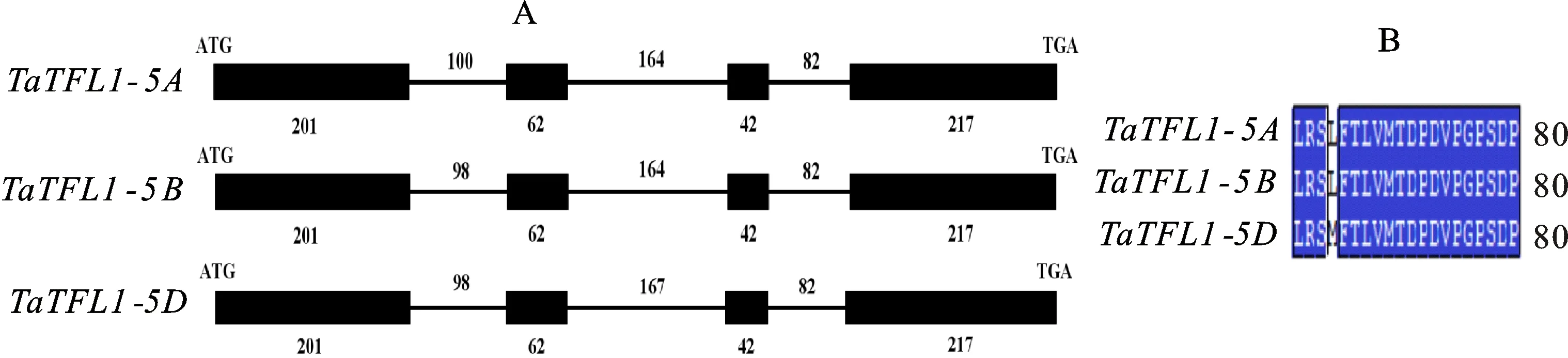
黑色方框代表PEBP家族典型的氨基酸结构和TFL1独特的氨基酸残基;黑色粗线条部分为TFL1同源蛋白底物结合保守域;Ta:小麦;Br:芜菁;Fc:无花果;Vu:豇豆;Hv:大麦;Lp:黑麦草;Tu:乌拉尔图小麦;Gm:大豆;Aet:山羊草;Bd:二穗短柄草;Os:水稻;Ob:短花药野生稻;TFL1和ZCN3分别来自拟南芥和玉米。Black box region represents a typical amino acid structure of PEBP gene family and unique amino acid residues of TFL1; the thick black line represents substrate binding conservative domain of TFL1 homologous protein; Ta:Triticum aeativum L.; Br:Brassica rapa; Fc:Ficus carica; Vu:Vigna unguiculate; Hv:Hordeum vulgare subsp; Lp:Lolium prernne; Tu:Triticum urartu; Gm:Glycine max; Aet:Aegilops tauschii; Bd:Brachypodium distachyon; Os:Oryza sativa; Ob:Oryza brachuantha; TFL1 and ZCN3 are derived from Arabidopsis thaliana and Zea mays,respectively.
2.3 TaTFL1蛋白多重比对及进化分析
用DNAMAN 5.0软件将乌拉尔图小麦(EMS48180.1)、黑麦草(AAG31808.1)、青稞(ABF85670.1)、玉米(ABX11005.1)、山羊草(XP_020199337.1)、二穗短柄草(XP_003578901.1)、水稻(XP_015617887.1 )、短花药野生稻(XP_006662737.1)等植物的TFL1同源蛋白与TaTFL1-5A(ATI23645.1)、 TaTFL1-5B(ATI23646.1)、 TaTFL1-5D(ATI23647.1)蛋白进行多重比对(图3)。从图3中可以看出,TaTFL1蛋白与其他物种的TFL1同源蛋白均具有PEBP家族典型的氨基酸结构:如70~74残基间的D-P-D-V-P模块及116~119之间的G-I-H-R模块,而且还具有TFL1亚家族独特的His88和Asp144残基,此结果进一步证实所获得的 TaTFL1基因序列的正确性。TaTFL1蛋白与其他单子叶植物的TFL1同源蛋白进行比对,其相似度为91.9%~100%,其中TaTFL1-5A和TaTFL1-5B蛋白与乌拉尔图小麦的同源蛋白一致, TaTFL1-5D蛋白与山羊草的一致,该结果表明,TFL1蛋白在单子叶植物中具有高度的保守性。而与双子叶植物比对的相似度也达到了69.27%~79.19%,可知TFL1蛋白在单子叶和双子叶中存在较大的差异。
利用MEGA 6.0软件,采用NJ法构建进化树,对TFL1蛋白在各物种中的进化关系进行了分析,结果(图4)表明,单子叶与双子叶植物的TFL1同源蛋白分别聚为两类,本研究得到的TaTFL1蛋白与乌拉尔图小麦和山羊草的亲缘关系最近,而与大豆和豇豆的亲缘关系最远。
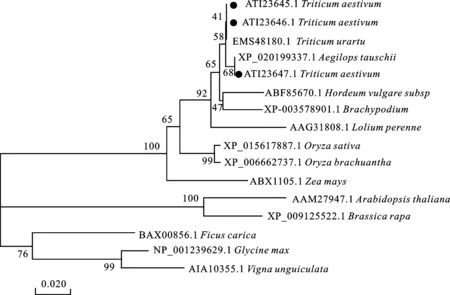
●代表本研究中 TaTFL1基因的编码蛋白。● indicates the deduced proteins of three TaTFL1 genes obtained in this study.
2.4 启动子的克隆及预测分析
以CS的gDNA为模板,以TP-F/TP-R为引物克隆得到 TaTFL1基因的上游序列,电泳结果如图1B所示。测序结果表明,共获得三类序列,进一步与Ensembl plants数据库比对,确认其分别为 TaTFL1-5A、 TaTFL1-5B和 TaTFL1-5D的上游序列。对启动子区进行预测,发现起始密码子上游1 200 bp的序列为启动子区。利用PlantCARE分析其中的顺式作用元件,结果发现这三个部分同源基因启动子的顺式作用元件组成完全一致,均含有多种参与光响应的元件,如AE-box、AT1-motif、Box 4、G-Box、G-box、Gap-box和sp1等元件,同时还含有与分生组织表达相关的顺式调控元件(CAT-box)、MYB转录因子结合位点(MBS)、赤霉素响应元件(P-box)、及与节律调控有关的顺式调控元件(circadian)(图5)。

图5 TaTFL1-5A、 TaTFL1-5B和 TaTFL1-5D启动子序列比对及其预测的元件分析
2.5 TaTFL1基因在中国春不同组织及不同时期的表达分析
不同组织中 TaTFL1的表达分析表明, TaTFL1在不同组织不同时期均有表达,但表达量存在差异。相对于其他组织,穗下茎中的表达量最高,且随着植株的逐渐发育,其表达量呈下降趋势(图6A)。在叶片中, TaTFL1有较低水平的表达,在刚萌发的芽中就开始表达,在幼苗期和花前时期叶片中的表达量明显高于花后灌浆期的表达量(图6B)。而在幼穗中,其表达量仅次于穗下茎中的表达量,随着幼穗的逐渐生长,其表达量逐渐降低(图6C)。颖壳中 TaTFL1的表达量无论在花期还是在花后都始终维持在很低的水平(图6D)。籽粒中的表达量总体较低,且随着籽粒的逐渐成熟而有明显的下降趋势(图6D),在幼根中也有低水平的表达(数据未列出)。由此可知, TaTFL1在小麦中的表达具有明显的组织特异性,且主要在开花前期的幼穗与穗下茎中表达。
3 讨 论
本试验对TaTFL1蛋白的功能进行预测,结果表明该蛋白行使转录调控功能的可能性最大,而前人的研究结果也表明了该基因具有转录调控功能。拟南芥和其他植物中的 TFL1基因过表达导致了营养生长延长,次级花序的增加,以及转基因拟南芥的开花延迟[15-19]。如在水稻中,过表达水稻的 TFL1同源基因导致水稻穗分枝增加,开花数是野生型的3倍,小穗排列更紧密,同时也引起了植株营养生长期延长,说明水稻中也存在着对 TFL1功能的响应途径[4];将黑麦草 LpTFL1转入拟南芥中过表达,植株开花明显延迟,侧分枝大量增加[9];将玉米的 ZCN1到 ZCN6过表达,发现同源性越高则表型表现得越强烈,能导致玉米晚花,花序的分枝大量增加[20];然而,将大麦 HvTFL1转入水稻中,其转化植株并未在开花时间和穗部结构发生改变,推测是大麦与水稻可能有着不同的开花途径[21]。由此可知, TFL1及其同源基因的功能在许多植物中是保守的,因此我们推测小麦中 TaTFL1可能也在维持花序分生组织状态和抑制开花的功能上存在相似性。
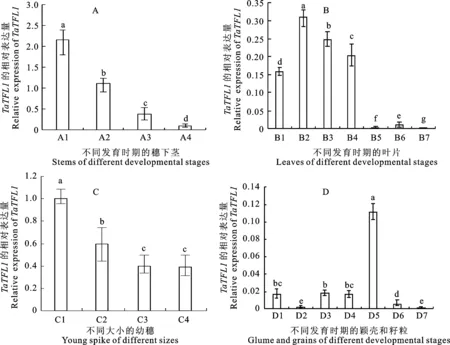
图柱上不同字母表示差异显著(P<0.05)。Different lower-case letters above bars indicate significant difference at 0.05 level.A1:花前;A2:花后5天;A3:花后10天;A4:花后15天;B1:芽;B2:萌发5天的叶;B3:萌发10天的叶;B4:花前的旗叶;B5:花后5天的旗叶;B6:花后10天的旗叶;B7:花后15天的旗叶;C1:1~2 cm幼穗;C2:2~4 cm幼穗;C3:4~5 cm幼穗;C4:5~6 cm幼穗。D1:花前的颖壳;D2:花后5天的颖壳;D3:花后10天的颖壳;D4:花后15天的颖壳;D5:花后5天的籽粒;D6:花后10天的籽粒;D7:花后15天的籽粒。A1:Pre-anthesis; A2:5 days after flowering;A3:10 days after flowering;A4:15 days after flowering; B1:Bud; B2:Leaf at 5 days after germination;B3:Leaf at 10 days after germination; B4:Flag leaf pre-anthesis; B5:Flag leaf at 5 days after flowering; B6:Flag leaf at 10 days after flowering;B7:Flag leaf at 15 days after flowering;C1:1-2 cm young spike; C2:2-4 cm young spike; C3:4-5 cm young spike;C4:5-6 cm young spike;D1:Glume pre-anthesis; D2:Glume at 5 days after flowering; D3:Glume at 10 days after flowering; D4:Glume at 15 days after flowering; D5:Grain at 5 days after flowering; D6:Grain at 10 days after flowering; D7:Grain at 15 days after flowering.
本研究克隆得到小麦第五同源群上的 TaTFL1部分同源基因,并测定 TaTFL1在不同组织不同时期的表达水平。结果发现小麦 TaTFL1-5A、 TaTFL1-5B和 TaTFL1-5D部分同源基因与拟南芥中 TFL1基因结构相同,同样由4个外显子和3个内含子组成[22]。本研究获得的3个 TaTFL1部分同源基因的CDS序列只存在较少的碱基差异,其预测的蛋白质表现为 TaTFL1-5A与TaTFL1-5B蛋白完全一致,而TaTFL1-5D与前者只在第64位氨基酸残基存在差异。本研究获得的3条 TaTFL1基因所编码蛋白的第88位氨基酸残基皆为His,且还具有TFL1家族典型的氨基酸结构,有研究表明TFL1蛋白的His88对其结构与活性具有至关重要的作用[19,23]。
TFL1基因虽然与FT基因的同源性较高,但两者的表达模式具有明显的差异。FT基因在叶片中的转录水平最高,其翻译的蛋白质作为成花素,从叶片中由韧皮部转运到茎顶端分生组织处,活化花分生组织特性基因的表达,从而促进开花;而 TFL1基因在顶端分生组织中的表达量最高[24]。在本试验中,小麦 TaTFL1在叶片中的表达量极低,结果与黑麦草[9]等相同,而与甘蔗[25]在顶端周围叶片中大量表达不同。Ratcliffe等[26]的研究结果表明,过表达 TFL1基因的拟南芥植株能产生更好的种子,也有人认为拟南芥中 TFL1能引起胚乳细胞化从而控制籽粒的大小。而在本试验中, TaTFL1在籽粒中的表达量较低,且随着籽粒的逐渐成熟而呈现出下降趋势,说明小麦 TaTFL1与拟南芥中 TFL1在籽粒发育中发挥的作用可能存在差异。为了明晰 TaTFL1是否参与了小麦幼穗的发育,本研究分析了 TaTFL1基因在不同大小幼穗中的表达情况,分析结果表明该基因在幼穗中的表达量较高,且随着幼穗的逐渐长大而表现出下降的趋势,推测小麦 TaTFL1基因主要在小麦幼穗发育初期起作用,该结果与拟南芥[27]、黑麦草[9]等植物中的研究结果相同,而且 TaTFL1启动子元件分析结果也表明其启动子中存在着分生组织表达相关的顺式调控元件(CAT-box)。本试验还发现 TaTFL1在穗下茎中大量表达,这与玉米中 ZCN1、ZCN2和 ZCN3在维管束中大量存在可能类似,具体原因还有待进一步的研究[20]。
