湖北松滋地区下奥陶统叠层石沉积特征
2018-10-08肖传桃吴彭珊李沫汝杨志伟周思宇郭瑞张超
肖传桃,吴彭珊,李沫汝,杨志伟,周思宇,郭瑞,张超
长江大学地球科学学院,武汉 430100
0 引言
自Kalkowsky[1]1908年提出“叠层石(stromatolite)”术语以来,人们对叠层石的认识在不断的深化和发展[2-5]。长期以来,在有关叠层石与微生物席、叠层石的形成机理、叠层石与矿产资源以及叠层石的分类和命名和其生物地层学意义等研究领域中取得了许多成果[6-23]。作为地球上最古老的化石之一,在地球历史上最早期的85%的记录中,叠层石是碳酸盐岩中最具有特色的沉积,是地球上探索生命起源和生物演化的直接证据[24]。因此,对叠层石形成的主控因素研究具有深入探讨生命起源和生物演化的重要意义。
叠层石是微生物岩中的一种最常见类型,是蓝细菌等底栖微生物生命活动与沉积作用相互作用而形成的一种特殊的原生纹层状、波状、柱状等生物沉积构造,这些原生生物记录了大量的微生物与环境、地球化学和地球物理等方面的信息[25]。早古生代的两次重要生物演化事件(寒武纪生物大爆发与奥陶纪生物大辐射)都与包括叠层石等微生物岩在内的生物礁生态系统密切相关[26]。生物大爆发后不久的寒武纪早期,以古杯类(Archaeocyathids)为主要格架的生物礁与微生物岩共生并大量繁盛[27-29]。寒武纪第二世末期的生物灭绝事件,导致古杯类动物造礁生态系统的全面崩溃[30-31]。这之后的寒武纪中晚期(即寒武纪第二世末期的生物灭绝事件到奥陶纪早—中期的生物大辐射事件之间约22个百万年的时间内),古海洋中动物骨骼较为匮乏,取而代之的是包括叠层石在内的大量微生物岩的繁盛。该时期一直被认为是微生物岩广泛发育的时期[32-34],也是生物礁生态系统由微生物岩建造向后生动物建造的转折时期[31],而该时期的海洋也被称为“贫乏骨骼的海洋”[35-36]。叠层石等微生物岩的繁盛与这些重大生物事件可能具有密切的联系,因此,详细查明叠层石等微生物岩的类型并深入进行研究,对理解这些重大生物事件具有一定的指导意义。
在已有的文献和成果中,有关前寒武系、寒武系、古近系、新近系和第四系叠层石研究成果较丰富,但关于奥陶纪叠层石方面的研究成果与实例很少[37-39];鉴于此因,作者认为,对下奥陶统叠层石的研究不仅丰富了我国叠层石的产出层位和类型、对于奥陶系烃源岩的勘探具有经济意义,更重要的是本研究成果对于探索微生物岩发育与中奥陶世后生生物大辐射之间的关系具有重要的科学价值。
1 区域地质概况
本次研究的叠层石发现于湖北松滋南部地区下奥陶统,该区位于江汉盆地的西南部,在大地构造位置上位于中扬子地台和江汉盆地的南缘。研究区奥陶系三统齐全,其地层序列与长江三峡地区一致,下统自下而上包括南津关组、分乡组和红花园组,中统包括大湾组和牯牛潭组,上统包括大田坝组、宝塔组和临湘组,但缺失五峰组。本区奥陶系以碳酸盐岩沉积为主,间夹页岩或钙质页岩,总厚约为463 m。在区域构造上,研究区奥陶系围绕以三叠系为核部的仁和坪向斜周缘、以寒武系为核部的刘家场背斜周缘分布,在奥陶纪时期研究区经历了局限台地、开阔台地、沉没台地、广海陆棚以及奥陶纪末期深水盆地环境的演化过程[40-41]。其中,南津关组、分乡组和红花园期均发育叠层石,形成了本区下奥陶统最具特征的微生物岩沉积。
本次发现的叠层石微生物岩主要见于研究区南津关组、分乡组和红花园组。南津关组下部为灰色厚层—块状亮晶砂屑、砾屑灰岩、凝块石灰岩、叠层石灰岩及条带状泥晶灰岩夹黄灰色页岩,厚76.5 m;上部为灰色中—厚层、厚层—块状亮晶砂屑、砾屑灰岩、泥晶白云岩、灰质白云岩夹鲕粒灰岩及白云质灰岩夹凝块石云质灰岩,厚97.8 m。南津关组产有较多的化石,按它们产出的层位特征,似可明显地划分为两个组合;下部为DactylocephalusdactyloidesAsaphopsisgranulatus组合,上部为Szechuanellaszechuanensis-Pseudobasilicussp. 组合。前者为宜昌剖面南津关组下部D.dactyloides-A.inflatus带的化石,后者亦为我国扬子区南津关组下部的重要分子,在长阳花桥,松滋卸甲坪均产于D.dactyloides带的上部层位。Asphopsis一属在西欧、北美均为Tremadocian阶下部的重要化石,因此,本区南津关组与宜昌地区南津关组可对比,其时代属于Tremadocian早期[37,40-41]。
分乡组下部主要为灰色中层亮晶砂屑灰岩、核形石砾屑灰岩夹凝块石灰岩等;上部为泥晶生屑灰岩、亮晶藻屑灰岩、薄层瘤状灰岩、厚层叠层石灰岩等与黄褐色页岩互层,化石丰富,本组厚71 m。分乡组产较多化石,所产的三叶虫Tungtzeellaszechuanensis、T. sp.为扬子区分乡组及其相当层位特有的带化石,分布很广。笔石化石较重要的有Acanthograptussinensis、A.sinensisvar.fenhsiangensis,其中Acanthograptus广泛分布于云、贵、川等地的桐梓组上部,为分乡组代表性带化石。牙形石较重要的有Drepanodusarcuatus、Paltodusdelifer、Paroistodusnumarcuatus等,其中Paltodusdeltifer是始于分乡组之底、消失于分乡组之顶的重要化石,Drepanodusarcuatus为分乡组P.deltifer-Triangulodusproteus带的重要分子(朱忠德等,1995)[37,40-41]。Paroistodusnumarcuatus则为欧洲Tremadocian阶上部重要化石,因此,本区分乡组的层位应属于Tremadocian晚期。
2 叠层石发育特征
研究区的叠层石主要分布于松滋刘家场地区的南津关组、分乡组和红花园组(图1),从叠层石的发育演替情况来看,构成了两个叠层石建礁旋回,南津关组为第一个旋回,分乡组和红花园组为第二个旋回。
2.1 纵向发育特征
在上述三个叠层石建礁旋回中,南津关组发育3层叠层石,分乡组和红花园组各发育一层叠层石。总体来讲,研究区下奥陶统自下而上叠层石的层数、厚度逐渐减少,规模也向上逐渐变小,但三个组沉积时期有各自的个性。其中,南津关组和分乡组规模最大,红花园组规模较小。在南津关组内部,第一层和第二层叠层石厚度和规模较大,第三层叠层石规模较小,形成了规模由大至小的代表下奥陶统第一个叠层石建礁旋回。分乡组和红花园组各发育一层叠层石,不同的是分乡组叠层石厚度和规模很大,红花园组叠层石规模较小,两者构成了一个由大变小的叠层石建礁旋回。
2.2 横向发育特征
从横向发育特征来看,南津关组第一层叠层石发育在第二段的底部,为灰色厚层—块状叠层石灰岩,厚约2.7~3.2 m。其底部为浅灰—灰白色色亮晶生屑含砾砂屑灰岩,顶部为灰色、黄灰色泥质条带灰岩。本层叠层石在横向上的发育特征变化较大,不仅其层厚发生一定的变化,而且其纹层的叠置样式和内部结构也有明显变化,在洈水响水洞一带自东而西叠层石类型呈现出柱状、大型波状、纹层状以及凝块状(详见下文中讨论部分),呈现叠层石类型变化的横向露头范围大约为2 km左右。在凝块状叠层石顶部可见食草动物腹足类Ecculiomphalus等生物化石。南津关组第二层叠层石发育在第二段的下部,为灰色厚层—块状叠层石灰岩,厚约2.5~2.9 m,其底部为浅灰色亮晶砂屑灰岩,顶部为黄色钙质泥岩。与第一层叠层石类似,本层叠层石在横向上的发育特征变化也较大,其纹层的叠置样式和内部结构也呈现出有规律的变化,在研究区自东而西叠层石的类型由柱状、大型波状变为纹层状,其中柱状叠层石不仅单独成礁,而且和Pelmatozoan(有柄棘皮动物)联合建礁,形成了地史上最早的Pelmatozoan生物礁[39]。南津关组第三层叠层石发育在第四段,为深灰色中—厚层叠层白云岩,厚约2.5~2.9 m,其底部为浅灰色亮晶砾屑灰岩,顶部为灰色中层亮晶砂屑灰岩。与前两层叠层石不同,本层叠层石在横向上的发育较局限,体现为一个规模较小的丘状叠层石。
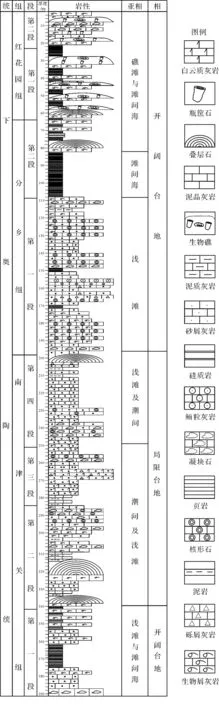
图1 松滋地区下奥陶统岩性柱状图Fig.1 Lithologic column of Lower Ordovician in Songzi area
分乡组叠层石发育在第二段下部,为灰色厚层—块状叠层石灰岩,厚约7.5~8.2 m,其底部为浅灰色亮晶砂屑、生屑灰岩,顶部为黄色泥岩。本层叠层石在横向上也具有不同的发育特征,其纹层的叠置样式和内部结构呈现出有规律的变化,在研究区响水洞至雷家塌一带自东而西叠层石的类型由柱状、大型波状变为纹层状,其中柱状叠层石不仅单独成礁,而且和Calathium(瓶筐虫)联合建礁,形成了地史上最早的Calathium生物礁[42-43]。
红花园组叠层石发育在本组底部,为灰色厚层—块状叠层石Calathium灰岩,厚约2.2~2.5 m,其底部为浅灰色亮晶生屑灰岩,顶部为黄色泥岩。本层叠层石主要以藻纹层形成存在,可以呈现独自互相包裹的纹层,或者围绕Calathium包裹生长等形式生长。在横向上其发育特征变化较小。红花园组叠层石主要集中于下部,向上明显减少。
3 叠层石的类型及沉积特征
有关叠层石的分类方案较为复杂,目前的分类方法主要有四种,一是根据叠层石的成因进行分类,分为骨骼叠层石、黏结叠层石、石灰华叠层石、细粒叠层石和陆生叠层石等;二是根据叠层石的形态类型进行分类,并分为超群、群、形、亚形等级别;三是根据生物学采用的双名法对叠层石进行属、种的分类[14,21],但属、种不具有生物学演化上的含义;四是根据叠层石几何形态进行分类,可以分为:层状叠层石、波状叠层石、丘状叠层石、柱状叠层石、指状叠层石等。关于叠层石的分类述评有学者已经进行了讨论[44],本文不做赘述。由于前人对奥陶纪叠层石研究较少,本文根据几何形态特征将研究区的叠层石分为以下几种类型,即层状、波状、柱状、丘状以及凝块石状,其中凝块状叠层石是本文新命名的叠层石类型。
3.1 层状叠层石
层状叠层石又称席状叠层石,其纹层具有近水平或略有波状起伏特征(图2A)。研究区该类层状叠层石较为发育,在南津关组,分乡组中均有分布,叠层石灰岩外观呈层状至块状,其纹层近水平展布,略有小的波状起伏,岩层的整体厚度和形态而无明显变化。在野外风化面上层状叠层石纹层较粗糙、欠规则,纹层之间可见少量生物碎屑和砂屑。南津关组层状叠层石局部发育微小的不规则凝块,凝块大小小于1~2 mm。暗纹层为主体表现为深灰色不规则条带,亮纹层表现为灰色断断续续不规则条带,纹层明暗间隔明显,总体上亮层要大于暗层,纹层对厚约7~15 mm,暗层1~5 mm。层状叠层石灰岩单层厚度不稳定,厚约0.4~3.6 m不等,但分乡组略厚于南津关组。在显微岩相特征方面,层状叠层石具有不均一的结构,总体来讲,叠层石灰岩以泥晶方解石为主要组成,其次可见少量生物碎屑,如棘皮类和腕足类碎屑,还可以发育少量砂屑。局部暗色纹层横向变为暗色细小的凝块,凝块也由泥晶团块组成。叠层石的暗层中可见大量的Girvanella丝状体微生物(图3D),在暗层中呈互相缠绕状,且大致平行于纹层展布,其周围多为泥晶方解石,也可见白云石化晶体的存在。
3.2 波状叠层石
波状叠层石是指纹层具有波状起伏特征的叠层石,该类叠层石在研究区最为发育,在南津关组、分乡组和红花园组中均有分布(图2B,C,D)。其中,南津关组和分乡组中的波状叠层石灰岩外观呈层状,且横向上分布于层状叠层石与柱状叠层石之间,其纹层多表现为较为规则起伏的波状,波长为5~40 cm,以15 cm左右为主,波高为1~15 cm,以5 cm左右为主;红花园组叠层石则多呈块状或丘状,其纹层不太规则,介于波状和柱状之间,波长约6~10 cm,波高5~15 cm。而红花园组叠层石呈不规则波状,其波峰高低起伏不平,纹层较清晰,多呈包裹状。在野外风化面上叠层石纹层较粗糙,且纹层厚薄不一,粗细不均,纹层之间可见少量生物碎屑和砂屑。横向上看南津关组中相邻波状纹层之间的波谷处可见有少量的凝块,大小约1.5~4 cm不等,此外,暗色纹层在横向上延伸中局部相变为微小的凝块,凝块大小1~2 mm。暗纹层为主体表现为深灰色不规则条带,亮纹层表现为灰色断断续续不规则条带,纹层明暗间隔明显,总体上亮层略大于暗层,纹层对厚约6~19 mm,暗层0.5~5 mm。波状叠层石灰岩单层厚度不稳定,厚约0.5~4.9 m不等。

图3 松滋刘家场地区下奥陶统叠层石特征(2)A.柱状叠层石,a为柱状双锥叠层石,产自南津关组;B.凝块状叠层石,白色箭头为凝块,朝上的黑色箭头为纹层,朝下的黑色箭头为纹层放大,产自南津关组;C.凝块状叠层石中的钻孔动物腹足类Ecculiomphalus,白色箭头为Ecculiomphalus,黑色箭头为凝块,产自分乡组;D.层状叠层石显微特征,箭头为Girvanella管状体相互缠绕形成的暗纹层,产自南津关组;E.波状叠层石显微照片,箭头为Girvanella管状体斜交于亮纹层中,产自红花园组;F.叠层石中纹层之间的棘皮碎屑,产自南津关组Fig.3 Characteristics of stromatolites of Lower Ordovician in Songzi area(2)
在显微岩相特征方面,波状叠层石具有不均一的内部结构,横向上,波状叠层石分为波状纹层系列和波状纹层间,纵向上,可分为纹层与纹层间。纹层间以泥晶方解石为其主要组成,其次可见少量生物碎屑和砂屑,生物屑如棘皮类,局部暗色纹层横向变为暗色细小的凝块,凝块也由泥晶团块组成。相邻的波状纹层间构成了波状叠层石特征的微相—水道间沉积,它代表了波状叠层石生物礁礁间水道微相,主要充填生物碎屑和砂屑,生物屑以海百合茎干为主,其次为由泥晶团块组成的凝块。叠层石的暗层中可见大量的Girvanella丝状体微生物,在亮纹层中呈斜交或垂直纹层方向发育,其周围为泥晶方解石、部分已经重结晶(图3F)。该类叠层石构筑于亮晶含砾砂屑灰岩之上,分乡组发育于亮晶生物屑之上。
3.3 柱状叠层石
柱状叠层石是指外观呈柱状形态的叠层石,该类叠层石在研究区较为发育,主要见于南津关组和分乡组(图2F、图3A),叠层石灰岩外观呈厚层状至块状,且横向上有序分布于研究区波状叠层石与层状叠层石的东侧。柱状叠层石由相互独立的垂向上堆叠的、向上弯曲或上凸的许多柱状体组成,每个柱状体内部由暗色藻纹层和浅色钙质沉积薄层向上相间排列叠置而成,柱状体内部四周纹层往往会下垂弯曲而形成外壁。柱状叠层石的柱体横断面多呈圆形、椭圆形及不规则椭圆形等,直径8~50 cm不等,高约40~180 cm不等。叠层石的柱状体纵切面呈不同的形态,有的呈下粗上细,有的呈上粗下细,有的下粗上尖,有的呈中间粗上下两头尖的,根据横断面及纵切面的形态组合特征,研究区的南津关组和分乡组柱状叠层石可以分为圆柱状叠层石(下粗上细)、锥柱状叠层石(下粗上尖)、倒锥柱状叠层石(上粗下细)和双锥柱状叠层石(中间粗上下两头尖的,新命名),其中前三种类型见于豫西寒武系馒头组中[19],双锥柱状叠层石为本区首次发现的新类型(图3A)。在柱状叠层石的各柱体之间充填的沉积物均为生物碎屑和砂屑,生物碎屑的类型主要为有柄棘皮动物的茎干碎屑,含量大约为40%左右,其次为少量砂屑。柱状叠层石的纹层对厚约8 mm,亮层发育,几乎相当于纹层对厚度,但暗层极不发育,约厚0.1 mm左右,较波状叠层石和层状叠层石薄很多,此种现象可能与蓝细菌不太发育有关。值得指出的是,在部分柱状叠层石内部可见有柄棘皮动物的原地垂直固着生长,他们与蓝细菌一起形成障积—黏结礁[19]。
在显微岩相特征方面,柱状叠层石也具有不均一的内部结构,横向上,柱状叠层石分为柱状体和柱体间两部分,纵向上也可分为纹层与纹层间。纹层间以泥晶方解石为其主要组成,其次可见生物碎屑和砂屑,生物屑如棘皮类(图3F),较少见凝块。相邻的柱体间构成了柱状叠层石特征的微相—礁间水道沉积,其规模比波状叠层石间的水道大,水动力更强,代表了柱状叠层石生物礁礁间水道微相,主要充填生物碎屑和少量砂屑,生物碎屑等均被亮晶胶结物充填。叠层石的暗色纹层不太发育,其中偶见葛万菌类丝状体微生物,纹层间周围多为亮晶方解石、部分为泥晶方解石。该类叠层石多半构筑于亮晶生屑灰岩之上。
3.4 丘状叠层石
丘状叠层石是指形态上向顶部方向凸起而无向下凸出的半圆丘状叠层石,该类叠层石在研究区不太发育,少量见于南津关组二段和分乡组二段,且多半独立发育或者发育于层状叠层石的之间(图2E)。其纹层延伸规律性较好,呈丘状弯曲的纹层往往会在叠层石的边缘融合,没有向底部方向的凸出,南津关组叠层石纹层较清晰,分乡组暗色纹层不发育。丘状叠层石宽为5~16 cm,高为10~30 cm。南津关组纹层内部的基本层微构造一般为断续的凝块定向排列而成。在显微岩相特征方面,丘状叠层石也具有不均一的内部结构,横向上,相变为砂屑砂屑灰岩或者层状叠层石灰岩。纵向上也可分为纹层与纹层间。纹层间以泥晶方解石为其主要组成,生物碎屑或砂屑较少。该类叠层石构筑于的砂屑、生屑灰岩之上。
3.5 凝块状叠层石
凝块状叠层石是本文新命名的一种叠层石,其形态上呈凝块状、不规则状断续或连续杂乱分布(图3B)。大型结构呈层状,但中型结构与凝块石相似,很难区别,如果仅根据其外表特征可以鉴定为凝块石,但从野外横向延伸上追索发现凝块状叠层石与层状或波状叠层石相连,故命名为凝块状叠层石。该类叠层石在研究区发育较少,仅见于南津关组二段第一层含叠层石地层中。凝块状叠层石灰岩外观呈厚层状至块状,侧向上厚薄不一,且横向上分布于层状叠层石与波状叠层石之间。叠层石内部形态呈凝块状、不规则状断续或连续的杂乱分布,凝块风化面呈灰棕色,新鲜面呈深灰色,局部结成斑状,或不规则网状,局部隐约可见凝块呈现断续的纹层特征(图3B),但暗色层厚薄不一,纹层间隐约可见少量砂屑及生物屑。
在显微岩相特征方面,凝块叠层石也具有不均一的内部结构。准纹层间以泥晶方解石为其主要组成,其次可见少量的生物碎屑和砂屑。暗色层多表现为凝块,其内部结构与凝块石很相似,以泥晶团块为特色,其中偶见葛万菌类丝状体微生物,其周围多为泥晶方解石。该类叠层石多半构筑于亮晶砂屑灰岩之上。
值得指出的是在凝块叠层石的层面上可见食草动物腹足类Ecculiomphalus化石及生物扰动痕迹(图3C),认为研究区凝块叠层石是生物扰动造成的(见下文讨论)。
4 叠层石的沉积模式及环境特征
根据叠层石发育的形态类型,结合不同类型叠层石内部的岩性与沉积特征,对研究区各类叠层石沉积环境进行如下分析,并总结了其沉积模式(图4)。
4.1 层状叠层石
该类叠层石主要特征是呈水平层状或较低的波状形态,发育的岩性主要为泥晶灰岩及白云质灰岩,仅在暗色纹层间看见少量砂屑或粉屑。叠层石局部暗色纹层横向变为暗色细小的凝块,且凝块也由泥晶团块组成。此外,在该类叠层石灰岩中未见暴露沉积相标志,叠层石的暗层中可见大量的葛万菌类丝状体微生物,该类生物多半生活于水体不太开阔的水体中。叠层石底部发育亮晶砂屑灰岩,顶部局部含少量泥质。综合以上沉积特征认为,层状叠层石可能形成于潮间带环境中,可能生活于潮间带偏上部的生态环境。
4.2 波状叠层石、丘状叠层石与凝块状叠层石
波状叠层石在横向上分布于层状叠层石与柱状叠层石之间,其纹层多表现为较为规则起伏的波状,其发育的岩性主要为泥晶灰岩,其颜色呈灰色至深灰色。暗色纹层在横向上延伸中局部相变为微小的凝块,横向上看南津关组中相邻波状纹层之间的波谷处可见有较多的凝块,大小约1.5~6 cm不等。纹层的波谷处除少量凝块之外,还多见砂屑及生物屑,在暗色纹层间可见少量砂屑或粉屑。上述现象反应了波状叠层石形成需要一定的能量,应该较层状叠层石形成的能量稍高。叠层石灰岩被击打成碎块后臭味较浓,反映了微生物岩的有机质仍然保存较好,也反映了其水体较层状叠层石略深。叠层石的暗层中也可见葛万菌类丝状体微生物,该类生物多半生活于水体不太开阔的水体中。叠层石底部发育亮晶砂屑、或生物屑灰岩,顶部局部含少量泥质(南津关组),或者为黄灰色页岩(分乡组)。丘状叠层石在横向上分布较为局限,其形成机理与波状叠层石大致相似。凝块状叠层石属于生物扰动后形成的特殊叠层石,从外貌上看,其内部应该保留了部分波状叠层石的特征,因此,形成环境与波状叠层石相似。综合以上沉积特征认为,波状叠层石可能形成于具有较弱能量的潮间带环境中,可能生活于潮间带偏中部的生态环境中。
4.3 柱状叠层石
柱状叠层石且横向上有序分布于研究区波状叠层石与层状叠层石的东侧,其柱体横断面多呈圆形、椭圆形及不规则椭圆形等。柱状体纵切面呈不同的形态,有的呈下粗上细,有的呈上粗下细,有的下粗上尖,有的呈中间粗上下两头尖的,它们可以分为圆柱状叠层石(下粗上细)、锥柱状叠层石(下粗上尖)、倒锥柱状叠层石(上粗下细)和双锥柱状叠层石(中间粗上下两头尖的)。纹层间以泥晶方解石为其主要组成。在柱状叠层石的各柱体之间充填的沉积物均为生物碎屑和砂屑,生物碎屑的类型主要为有柄棘皮动物的茎干碎屑,含量大约为40%左右,其次为少量砂屑,其胶结物多为亮晶方解石,它们代表了柱状叠层石生物礁间的水道微相,该类叠层石多半构筑于亮晶生屑灰岩之上。上述现象反应了柱状叠层石形成需要较高的能量,应该较波状叠层石形成的能量高。综合以上沉积特征认为,波状叠层石可能形成于潮间带下部至潮下带上部的较高能量的环境中。
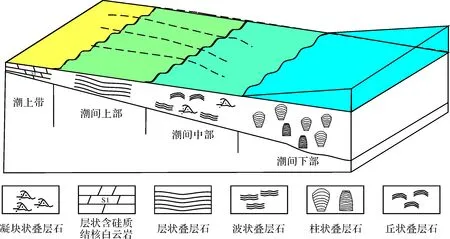
图4 松滋地区下奥陶统叠层石沉积模式Fig.4 Stromatolites Sedimentary Model sketch of Lower Ordovician in Songzi area
5 叠层石发育与中奥陶世后生动物大辐射之间的关系探讨
研究区下奥陶统时期存在三个叠层石发育旋回,即南津关组沉积期、分乡组沉积期和红花园组沉积期。从发育特征来讲,自下而上叠层石的层数、厚度逐渐减少,规模也向上逐渐变小,但三个沉积时期有各自的个性。其中,南津关期和分乡期规模最大,红花园期规模明显缩小并最终消失,从功能学来讲,下奥陶统的造叠层石微生物逐渐由建礁转为辅助建礁,并在弗洛期末期消失。研究区下奥陶统叠层石演化特征说明什么?究竟是什么原因导致这其在弗洛期末期消失?他们与壳体生物演化之间存在什么关系呢?这是许多学者正在思考的问题。
从生物的多样性研究来看,奥陶纪存在着三次生物演化的多样性高峰期[45],但最主要的是出现在早、中奥陶世的首次峰值。早、中奥陶世正是海洋无脊椎动物出现大分异、发生大辐射的重要时期。基于全球性的生物演化数据,Sepkoski对早古生代时期不同生物门类的多样性演化进行了分析[46],分析结果表明,尽管奥陶纪时期不同门类多样性发生了明显剧增,但它们辐射的起始时间有着明显的不同步现象。例如,三叶虫、腕足动物、笔石、大多数软体动物以及棘皮动物从奥陶纪初就已开始发生显著的多样性增长,而同样是营底栖固着、且以群体方式生活的苔藓虫与珊瑚的爆发始于大坪期(即阿仑尼格中晚期),而三叶虫演替体现为Whiterock 动物群替代Ibex动物群,其演替时间大致在晚奥陶世早期[47-49]。不仅如此,其他不同生物类群其多样性演化也具有相似的特征,如珊瑚几乎是呈指数型加速递增,而部分软体动物和腕足动物则以阶梯式递增[49]。浮游生态的笔石在中奥陶世早期就达到其多样性的顶峰。牙形石则在下奥陶统末几乎达到了顶峰期。但研究表明,奥陶纪最大的生物辐射是发生在中奥陶世大坪期[46]。因此,有些学者认为地史时期叠层石的逐渐减少、衰退与后生动物丰度增加有一定关系,换句话说,叠层石减少与后生动物不断啃食形成叠层石的微生物菌藻类有一定关系,如Reid 等在现代叠层石中发现大量内部纹层被蛤和海绵等生物破坏的痕迹,且有明显的生物痕迹被保存在纹层中[49];我国学者曹瑞骥等在我国湖北保康马桥神农架群石家冲组的叠层石中发现了生物钻孔和食草动物破坏留下的疤痕[14]。
总体来讲,本文赞成上述观点,即叠层石在纵向上逐渐减少的主要原因是后生动物大量增加并不断啃食形成叠层石的菌藻类微生物的结果。首先,这一点可以从生物演化的多样性特征得到证实,即下奥陶统末期及中奥陶世初期后生动物特别是底栖后生动物的大量增加与叠层石的减少有着对应关系;其次,在研究区的多数叠层石中我们发现了后生食草动物腹足类Ecculiomphalus化石(图3C),局部层段食草动物几乎呈集聚式存在,这些食草动物不仅啃食了形成叠层石的菌藻类微生物,造成叠层石数量的不断减少,而且破坏了叠层石的生长状态,并且形成了研究区独具特色的凝块状叠层石(图3B,C)。
此外,从中奥陶世开始,研究区海平面的快速上升也是叠层石减少的原因之一。由于海平面的较快速上升,研究区经历了由沉没台地、广海陆棚演化为奥陶纪末期深水盆地环境的过程[40-41],使得叠层石原有的生存环境因素和条件彻底破坏,从而也会导致中奥陶世开始叠层石快速衰减。因此,虽然后生动物大辐射是叠层石衰退原因之一,但环境条件的变化也是叠层石衰退不可忽视的因素之一。
6 结论
通过对松滋刘家场地区下奥陶统叠层石沉积特征的研究,取得如下认识:
(1) 研究区下奥陶统南津关组、分乡组和红花园组均发育叠层石,且叠层石在纵向上具有规模、厚度和数量逐渐减小的特征,在横向上其类型也发生有规律的变化,由水平层状、波状逐渐变为柱状的类型。
(2) 新发现并命名了凝块状叠层石和双锥柱状叠层石两种新类型。根据几何形态发育特征,本文将研究区的叠层石分为以下几种类型,即层状、波状、柱状、丘状以及凝块状叠层石。作者认为凝块状叠层石实际上是由于食草动物和扰动所形成的。
(3) 在对各类型叠层石的沉积特征进行详细阐述基础上,结合不同类型叠层石内部的岩性、微相及显微特征,对研究区各类叠层石沉积环境进行了分析,并总结了研究区叠层石的沉积模式。
(4) 对叠层石发育与中奥陶世后生动物大辐射之间的关系进行了探讨,根据中奥陶世初期后生动物的大量辐射与叠层石突然减少的对应关系,结合研究区叠层石中发现许多后生食草动物腹足类Ecculiomphalus化石等现象,认为叠层石随时间的逐渐减少、衰退与后生动物丰度增加有一定关系。指出食草动物不仅啃食了形成叠层石的菌藻类微生物,造成叠层石数量的不断减少,而且破坏了叠层石的生长状态,结果形成了研究区独具特征的凝块状叠层石。此外,研究认为,中奥陶世开始,海平面的快速上升也是研究区叠层石减少直至消失的原因之一。
