浙江省野生蜡梅花部形态变异及其与环境因子的相关性
2018-09-28杜会聪蒋雅婷王彩霞
杜会聪,蒋雅婷,田 敏,张 莹,王彩霞
中国林业科学研究院亚热带林业研究所,富阳 311400
1 材料与方法
1.1 研究地区自然概况
浙江省的野生蜡梅多生于避风、气候温和且湿润的低山沟谷及山脊两侧。特殊生境及人工破坏性的砍伐,使得年代较久的蜡梅数量极少,直接影响了野生群落的自然演替。目前,主要分布在杭州富阳区胥口镇的通天飞瀑风景区(TTFP)、洞桥镇上林村龙王坎(LWK)、洞桥镇石羊村碧东山(BDS)、万市镇白石村五尖山(WJS)和临安市玲珑镇高山村方山境内(FS),其地理位置如表1所示。碧东山和龙王坎的蜡梅多沿陡峭的溪谷呈狭带状分布,主要分布在山坡的中部和谷底,立地环境恶劣,蜡梅为获取足够光照,向溪谷侧斜向生长。通天飞瀑、方山和碧东山为常绿阔叶林,群落树冠连续分布,呈波浪状起伏,林相相对整齐,群落高度均在4.5—6.5 m,总郁闭度大约为90%,局部可达100%。伴生植物主要有:化香树(PlatycaryastrobilaceaSieb.et Zucc.)、浙江楠(Phoebechekiangensis)、枇杷(Eriobotryajaponica)、桂花(Osmanthusfragrans)、紫楠(Phoebesheareri)、糙叶树(Aphanantheaspera)、青冈(Cyclobalanopsisglauca)、枳椇(Hoveniaacerba)、络石(Trachelospermumjasminoides)、野菊(Dendranthemaindicum)等;五尖山分布区蜡梅数量较多且分布相对集中,该群落无乔木层,蜡梅是优势种,其他灌木和草本较少,郁闭度低;龙王坎分布区上层有稀疏乔木层,灌木和草本植物受其影响呈稀疏分布,林分郁闭度相对偏中,伴生植物主要有:柏木(Cupressusfunebris)、化香树、盐肤木(Rhuschinensis)等。部分群落外貌有藤蔓交错的现象,藤蔓植物主要有紫藤(Wisteriasinensis)、三叶木通(Akebiatrifoliata)、常春油麻藤(Mucunasempervirens)等,丰富的藤蔓植物客观上对野生蜡梅的生长、演替和保护具有一定的积极意义。

表1 各种群的地理位置与生境概况
LWK:Longwangkan; BDS:Bidongshan; TTFP:Tongtianfeipu; WJS:Wujianshan; FS:Fangshan
1.2 形态特征测定
在蜡梅盛花期,在各居群内随机选取9—15株生长正常、无严重缺陷和病虫害的蜡梅。为了避免采样株间的亲缘关系[10],株间距均在30 m以上。单株上随机采取位于向阳方向枝条中部的5—10朵蜡梅花,分层置于较大体积的容器运回并立即测量各指标,防止运输中挤压和花萎缩对花型和各参数的影响。统计中被片数、内被片数、雄蕊数、雌蕊数和胚珠数,测定中被片长、内被片长、中被片宽、内被片宽、花冠直径(花冠外轮直径最大值)、花冠内径(花冠内轮直径最大值)、筒深、雄蕊长和雌蕊长,计算中被片长宽比和内被片长宽比,各指标均用精度为0.01 cm的电子游标卡尺测定,共16个形态指标。
1.3 土壤养分因子测定
在每一样方内按照S型曲线选取取样点,挖取根系附近距地表20 cm左右的土壤样品,混合后自然风干、过筛备用。土壤pH值测定采用电位法,半微量凯氏法测定全氮含量,速效磷采用钼锑抗比色法;速效钾和Na离子采用火焰原子吸收法,钙(Ca)、镁(Mg)、铁(Fe)、锌(Zn)、铜(Cu)和锰(Mn)等矿质元素的测定采用原子吸收光谱法。由国家林业局经济林产品质量检验检测中心(杭州)完成。各样地土壤养分含量的测定结果见表2。

表2 浙江省蜡梅居群土壤养分特征
1.4 数据处理
使用Excel 2007、SPSS 20.0和SAS 11.5软件包提供的有关程序进行统计分析,计算各指标的算术平均值、标准差和变异系数;通过变异系数衡量表型性状及种群的变异水平;对各性状及其与环境因子的相关性进行分析;主成分分析得出表型性状对种群变异的贡献率;最后进行Q型聚类(UPGMA法)和R型聚类分析[11- 12]。
巢式方差[13]分析蜡梅性状变异的线性模型为
Yijk=μ+Pi+Ti(j)+e(ij)k,
式中,Yijk为第i群体第j个个体第k个观测值,μ为总均值,Pi为第i个群体的效应值(固定),Ti(j)为第i个群体内的第j个个体的效应值(随机),e(ij)k为试验误差。
以表型分化系数[14](Vst)反应群体间表型分化。

多洛肯,男,新疆霍城人,博士,教授,博士研究生导师,主要研究方向为古今少数民族汉文学、元明清文学古籍整理、元明清诗文。
变异系数计算公式:
CV(%)=S/x×100%
式中,CV为变异系数;S为标准差;x为平均值。
2 结果与分析
2.1 蜡梅居群花部的形态特征
野生蜡梅5个居群16个花部性状变异的测定结果显示(表3):不同的花部性状在群体间存在一定差异。花冠直径、筒深、中被片长、内被片长和雌蕊长均以碧东山居群的数值最大,其中花径、筒深、中被片长的数值极显著高于其他4个居群;临安方山居群的花冠直径、花冠内径、筒深、内被片长度和内被片长宽比数值最小,其中花冠直径、花冠内径和内被片长宽比的数值极显著低于其他4个居群;数量形状上,方山居群内被片数最多,雄蕊数、雌蕊数和胚珠数均值最小;通天飞瀑居群雌蕊数和胚珠数均值最大,中被片数和内被片数均值最小。在各居群间花冠直径的均值差异最大,碧东山均值为25.0789 mm,临安方山的均值为15.5073 mm;内被片个数、雄蕊长度、雌蕊数、雌蕊长度的均值在各居群间差异不显著。
2.2 蜡梅花部性状变异分析
变异系数可以反映蜡梅花部形态特征的变异程度,表3显示花部各性状间的变异程度各不相同,同一性状在不同居群间变异程度也不同,如筒深在不同居群间变异系数差值较大,五尖山居群的变异系数为0.3615,而通天飞瀑居群的变异系数仅为0.1220;同一居群不同性状变异系数也不相同,如通天飞瀑居群内胚珠数的变异系数是0.2396,而花冠内径的变异系数仅为0.0771,差距较大。在居群间五尖山居群变异系数均值最大为0.1765,其他四个居群数值相近。5个居群的16个花部性状中,变异系数均值从大到小依次为:胚珠数、内被片长度、筒深、雌蕊长度、雌蕊数、雄蕊长度、内被片长宽比、内被片宽度、花径、雄蕊数、中被片宽度、中被片长度、中被片长宽比、内被片数、中被片数、花冠内径(胚珠数的平均变异系数最大为0.2338,花冠内径最小为0.1081)。通过变异系数的分析可知, 蜡梅花部各个性状的变异程度在群体内和群体间有一定的差异,说明浙江省野生蜡梅有着丰富的形态变异。按巢式方差分析得到的各方差分量和表型分化系数可知:花部性状表型分化系数(Vst)的变异幅度为1.57%—89.62%,其中花冠直径、花冠内径、筒深、中被片长、中被片长宽比、内被片长、内被片长宽比、雄蕊数的表型分化系数在50%以上,说明这些性状在群体间的变异大于群体内变异,剩余与外界环境接触较少的花部性状的群体间变异小于种群内变异。16个性状的平均表型分化系数为44.38%,说明蜡梅花部形态变异以群体内的变异为主。
2.3 蜡梅花部性状的主成分分析
对蜡梅花部性状的数据进行主成分分析(表4)表明:前3个主成分方差累计贡献率为88.286%,可以代表原始性状的绝大部分信息。第一主成分方差贡献率为49.113%,起主导作用的性状是花冠直径、花冠内径、花筒深度、中被片长、内被片长,主要反映了花部花型。第二主成分的方差贡献率为22.981%,起主导作用的性状是中被片数。第三主成分方差贡献率是16.192%,起主导作用的性状是胚珠。由此说明与花大小相关的性状对变异起主要作用。

表3 浙江省5个蜡梅居群花部形态指标均值、变异系数及其显著性情况
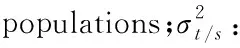

表4 蜡梅天然种群的主成分分析
2.4 相关性分析
2.4.1 花部性状间的相关性分析
植物在生长过程中各个形态特征间存在一定联系,相关性分析可以反映各性状间的影响关系。对5个种群16个性状相关性分析结果表明(表5):野生蜡梅花部各性状间呈一定的相关性,如中被片长与除中被片数和内被片数以外的其他性状均呈极显著相关关系,相关个数最多;内被片数与胚珠数有显著相关关系,与其他性状均无显著关系,相关个数最少。性状内部花冠直径、花冠内径、筒深、中被片长、中被片宽和内被片长这6个花外部性状间相互存在显著相关关系,其中花冠直径和中被片长度极显著相关(0.603),说明这6个性状间相互影响较大。数量性状中雄蕊数、雌蕊数、胚珠数三者相互间为极显著相关,中被片数和内被片数没有与任何性状呈极显著相关关系。
2.4.2 花部性状与地理-土壤因子间的相关性分析
对蜡梅16个花部性状的均值与居群的地理-土壤养分因子进行相关性分析,结果(表6)表明:铜含量与花冠直径、中被片数和中被片长宽比呈显著正相关,相关系数分别为0.937、0.918和0.941;铁含量与中被片长宽比呈显著正相关(r=0.915)。钙含量与胚珠数呈显著正相关(r=0.907)。海拔与内被片数极显著正相关(r=0.978),与中被片数呈正相关,与其他性状均为负相关关系,不显著;中被片宽、内被片数、内被片宽与经度呈正相关,中被片数、中被片长宽比、内被片数和内被片宽与纬度呈正相关;其他性状与经纬度呈负相关关系,均不显著。多数性状与土壤养分因子正相关,少数为负相关。计算大量元素、中量元素和微量元素与花性状的相关系数均值分别为0.422、0.327、0.413,由此说明大量元素与花部性状变异程度的相关性最大,微量元素次之,中量元素最弱。各环境因子对蜡梅花部表型性状的影响程度有一定差异,按照相关系数均值从大到小依次为土壤pH值、锌含量、磷含量、钠含量、经度、氮含量、纬度、铁含量、海拔、锰含量、钾含量、镁含量、铜含量、钙含量。

表5 野生蜡梅花部性状相关性分析
*:α=0.05水平上显著;**: α=0.01水平上显著

表6 蜡梅花部性状与地理土壤因子的相关系数
2.5 聚类分析
基于16项花的性状特征值对5个天然居群进行聚类分析,结果(图1)显示:在欧氏距离临近10处可以把5个居群可以分为2组。通天飞瀑居群与五尖山居群在最短截取处聚类,然后分别再与龙王坎居群和临安方山居群形成第一大组,第二组碧东山居群单独一组,其中通天飞瀑、五尖山居群遗传距离较小说明两者间的分化程度最小,龙王坎居群与方山和通天飞瀑居群、五尖山居群遗传距离相近,而碧东山与其他居群遗传距离最远,分化程度最大。
对蜡梅花部性状的R型聚类分析可知(图2),在遗传距离临近15的位置可以将蜡梅花部性状分为两大类。第一类为花冠直径、筒深、中被片长,包括了花外部形态相关的3大性状;第二类为除去上述三个性状的所有性状。在遗传距离5—10中间的位置则可以将蜡梅花部形状分为4大类,上述第一类分为A、B两类,A类包括中被片长、筒深,两者决定着蜡梅的花瓣的伸展角度,能体现其花型;花冠直径为一类为B类,花冠直径也是蜡梅分类标准中特别重要的一个性状。上述第二类分为C、D两类,C类主要为花部营养性状,其中包含中被片宽、内被片宽、中被片长宽比、内被片长宽比、雄蕊长、雌蕊长;D类主要为花部相关的数量性状,其中包括花冠内径、内被片长、中被片数、内被片数、雄蕊数、雌蕊数、胚珠数。

图1 基于花部性状分析结果的5个蜡梅居群UPGMA聚类图

图2 蜡梅花部形态特征性状R型聚类分析图
3 结论与讨论
3.1 蜡梅花部形态变异的多样性
表型性状是植物基因型和所处生态环境的综合体现[15]。对蜡梅5个居群花部性状多样性研究结果表明:碧东山居群的蜡梅花冠直径和中被片长的均值分别为25.0789 mm和16.3609 mm,显著大于国内其他野生居群[16-18]花冠直径和中被片长度的数值(花径和中被片最大均值分别为17.996 mm和14.056 mm[17])。由此说明浙江省野生蜡梅是优良的种质资源;各个居群内不同性状的变异系数存在较大的差异,变异范围是7.60%—36.15%,平均变异系数为15.03%,高于扇脉杓兰(Cypripediumjaponicum)[19](CV=11.143%)表型性状的平均变异系数,说明蜡梅存在丰富的遗传变异。野生蜡梅种群间平均表型分化系数为44.38%,高于玫瑰(Rosarugosa)[20](29.52%),低于张彩霞等[21]分析蜡梅群体8个性状的平均表型分化系数,这表明蜡梅天然群体间花部表型分化系数较大,可能存在较高的群体内自交率,调研中也发现授粉昆虫较少,这与蜡梅开花物候和生境海拔有关。与张彩霞等分析结果的差异可能是选取性状不同所导致。蜡梅居群花部形态变异以居群内为主,对于一个物种遗传结构来讲,植物本身的繁殖方式起决定作用[22]。Hamrick等[23]也指出,异花授粉物种的遗传变异主要发生在居群内,而自花授粉则相反。
3.2 野生蜡梅花部形态变异与环境因子关系
各环境因子与花性状的相关系数的均值中pH的均值最大,土壤酸碱度是土壤中化学性质的综合反映,土壤中各营养元素的转化、释放,镁、铁、钙等微量元素的有效性,有机质的合成与分解以及微生物的活动都与土壤酸碱度有关[24],且由土壤中养分因子和各性状的相关性可知,对性状形成绝对的支配作用需要多个养分因子甚至所有环境因子的综合作用,土壤酸度与其他单独的养分因子相比,综合性更强,因此对花部性状影响较大。花部性状与土壤中大量元素氮、磷、钾含量相关性最大,微量元素次之,中量元素最小,氮、磷、钾作为大量元素是植物体内如核酸、蛋白质、叶绿素等重要化合物的组成成分,也是土壤易缺乏的元素,能直接或者间接影响植物的生长发育和代谢;蜡梅花部的部分性状与土壤中的钙、铁和铜含量以及海拔有显著相关性,由此说明这4个因子的差异是形成蜡梅花性状多样性的原因之一,李根有等[5]在调研中也认为蜡梅属于喜钙树种。蜡梅花部各性状与环境-土壤养分因子的224对相关性组合中仅有6组显示有显著相关性,而茶条槭(Acerginnala)[25]17个表型性状与土壤中养分因子及地理坐标间有21组具有显著的正负相关性。蜡梅各居群土壤中氮和钾含量均值为235.17 mg/kg和90.98 mg/kg,均低于茶条槭(分别是388.08 mg/kg和113.8 mg/kg)和扇脉杓兰[19](分别是524.33 mg/kg和178.00 mg/kg)。浙江省野生蜡梅各居群生境的土壤类型多为黑色石灰土,部分蜡梅生活在河谷两侧陡坡甚至极薄的土层或石缝间,陡坡坡度在50°—80°较多,生存环境恶劣,土壤贫瘠。物种间的差异可能是土壤肥力、环境的异质性或者植物本身营养成分需求差异导致,当土壤满足植物生长所必需的营养成分时,物种各性状之间的差异较小。邓洪平[26]、李珊[9]、窦全丽[7]等分别进行了四川大头茶(Gordoniaacuminata)、云南金钱槭(Dipteroniadyeriana)、缙云卫矛(Euonymuschloranthoides)等种实的表性性状与土壤因子的相关性研究,证明了土壤因子对种实性状变异有一定的影响。环境因子包括很多内容,例如土壤厚度、透光率和其他未测元素含量等,本研究只调查了14个相关因子,这也是本研究的不足。
3.3 野生蜡梅保护策略以及待解决问题
蜡梅花颜色独特、芳香秀美,可提炼香精、入茶食用[27],且蜡梅在杭州栽培历史悠久[28],是城市园林绿化中应用较多的树种之一。作为华东唯一的野生蜡梅分布点,本地各居群遗传多样性水平较高,是蜡梅育种的重要基因资源。在野外调查发现该群落中的多数蜡梅正在经受病虫害困扰,存在大量腐烂枯枝,严重影响蜡梅的着花座果。群落生境条件差,且种子萌发困难,5个居群中仅在方山居群发现一些实生苗。实生苗最高在50 cm左右,多数小实生苗受人为干扰和环境影响不能存活,对群落的稳定性有很大影响。针对当前情况,建议采取以下保护策略,第一,对经受病虫害及生长势衰弱的蜡梅进行调查研究,找出原因进行治疗并修剪腐烂枯枝。对群落的演替进行一定的人为干涉,适当的疏伐,促进野生蜡梅的更新。第二,对蜡梅群落进行定期普查,蜡梅群落结构复杂,伴生种类繁多,结合蜡梅本身的适应性和人为干扰,建议至少2—3年对野生蜡梅资源进行普查,对其生长环境、生长势、保护现状进行监测。第三,建议在杭州富阳就近建立野生蜡梅基因库,保证其遗传多样性的原始性,制定有序的开发利用方案,有效保护和合理利用野生资源。
在调研中发现五尖山、龙王坎居群野生蜡梅开花量较大,但座果率极低甚至不到5%,远不及栽培蜡梅(70%左右),野生蜡梅开花物候较栽培蜡梅晚1个月左右正处冬季较冷的时段,且生境海拔高、土壤养分贫瘠,冬季温度低缺乏授粉昆虫或其他原因,致使蜡梅花部变异退化所致,具体原因将会进一步进行研究。在方山居群调研中发现了单株全部雄蕊瓣化的蜡梅花,无前一年遗留的果实。另有单株花型奇特,盛花期花冠口呈闭合或略开不到5 mm左右的小口如花苞状花型的蜡梅,这两种特殊蜡梅在浙江其他居群或蜡梅相关报道中均未出现,有待进一步观察研究。
