光周期对金银花叶片光合特性和抗氧化酶活性的影响
2018-09-12房海灵梁呈元
薛 欢,朱 梅,房海灵,梁呈元
(1.江苏省森林资源监测中心,江苏 南京 210036; 2.江苏省林业局,江苏 南京 210036; 3. 江苏省中国科学院植物研究所,江苏 南京 210014)
金银花为忍冬科植物忍冬(LonicerajaponicaThunb.),其干燥花蕾是我国重要的清热解毒类中药,广泛应用于医药、食品、化妆品等领域,市场需求量大[1-2]。人工种植金银花是满足当前原料市场需求的主要来源。金银花在我国已有较长栽培历史,在种植技术方面积累了丰富的经验,有效促进了金银花产业的发展。然而多数报道集中在对其田间管理、病虫害防治等方面[3~5],较少针对影响其生长发育的环境因子如光[6]、水[7]、肥[8]等进行研究。光(光强、光质和光周期)是影响植物生长发育的重要环境因子。赵倩[9]研究表明,短日照处理可使菊花初花期提前,盛花期可溶性蛋白、可溶性糖含量降低。董伟欣等[10]研究发现,短日照诱导能有效缩短小豆的生育进程(含花芽分化进程),其生育进程且随着诱导时间延长而加快,可见光周期对植物开花具有重要调控作用[11]。本研究通过采用人工遮光的方式,探究不同光周期处理对金银花叶片光合特征和抗氧化酶活性的影响,以期为人工栽培金银花中光照时间的确定提供依据。
1 材料与方法
1.1 材料
试验于江苏省中国科学院植物研究所试验基地进行,供试金银花品种为‘四季树型’,1年生苗。2015年11月下旬取生长一致金银花1年生扦插苗露地移栽,株行距1.0 m×1.0 m,常规水肥管理,备用。
1.2 试验处理
采用罩黑箱的方法人工控制光照时间。共设3个处理,即Ph 7:光照时长7 h(8:00—15:00),Ph 10:光照时长10 h(8:00—18:00),CK:自然光周期13 h(6:00—19:00,利用当地自然光照长度为对照,通常5月南京地区日光照长度13 h左右)。随机选取6株生长一致的金银花植株作为1个处理(3次重复)。试验于5月4日开始。处理开始时测量所有参试植株,株高约30.00—32.00 cm,地径 5.20—5.30 mm,1年生新梢生长量36.6—41.1cm。除光照时长外,所有处理均采用相同栽培管理措施进行管理。
1.3 检测项目及方法
1.3.1 光合色素含量测定 参考李合生[7]的方法略加修改。取已测定光合特性的金银花成熟叶片,去除叶脉后,用打孔器取直径0.5 cm叶圆片0.2 g,加25 mL 80%丙酮,避光浸提至叶片呈白色。将浸提液分别置于紫外分光光度计470,645,663 nm处测定吸光度。并根据公式计算光合色素含量:
叶绿素a质量浓度(mg/L)=12.21×A663 nm-2.81×A646 nm
叶绿素b质量浓度(mg/L)=20.13×A646 nm-5.03×A663 nm
类胡萝卜素质量浓度(mg/L)=(1 000×A470 nm-3.27×叶绿素a质量浓度-104×叶绿素b质量浓度)/229
根据以上色素计算结果换算成mg/g。
1.3.2 光合参数测定 在金银花二白期,于5月25日09:00—11:30利用LI-6400XT型光合仪测定生长一致的金银花新梢顶芽下第5—6对成熟叶片的光合特性。测定指标:净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr),并依据公式Ls=1-Ci/Ca(Ca为大气CO2浓度)计算气孔限制值。每个处理随机选取5株,重复叶片10片,计算平均值。
1.3.3 抗氧化酶活性测定 分别于光周期处理7,14,21,28,35 d时采集金银花新梢顶芽下第5—6对成熟叶片,测定其中3种抗氧化酶:超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)的活性。利用购自于南京建成生物技术有限公司的试剂盒完成。
1.4 数据统计与分析
试验所得数据采用Office Excel 2013和SPSS 19.0 软件进行统计和差异显著性分析,结果数据为平均值±标准误。
2 结果与分析
2.1 光周期处理对金银花叶片中光合色素含量的影响
由表1看出,光照时间的长短对金银花叶片中光合色素的合成具有显著影响,叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)含量随着光照时间的延长呈上升趋势,其中Chlb在光照时长10 h时增加幅度较大。Chla/b随光照时长增加呈先降低后升高趋势。
2.2 光周期处理对金银花叶片中光合参数的影响
光照时长对金银花成熟叶片光合参数具有显著影响。与CK相比,Ph 7和Ph 10处理可引起金银花叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)及蒸腾速率(Tr)显著下降。而气孔限制值则与其他指标呈相反变化趋势,即随光照时间缩短而上升。表明短日照处理可引起金银花叶片气孔导度减小,进而影响光合速率。
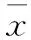
表1 光周期处理对金银花叶片中光合色素含量的影响±s (n=3)
同列数据后不同大、小写字母分别表示数据间存在极显著(P<0.01)、显著(P<0.05)水平差异
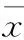
表2 光周期处理对金银花叶片光合参数的影响±s (n=10)
同列数据后不同大、小写字母分别表示数据间存在极显著(P<0.01)、显著(P<0.05)水平差异
2.3 光周期处理对金银花叶片中抗氧化酶活性的影响
2.3.1 SOD活性 与对照(CK)相比,在Ph 7和Ph 10处理组,金银花叶片中SOD活性表现出急速增加的趋势;处理28 d时,Ph 10处理组SOD活性低于CK组和Ph 7组,表明适度光照时间有效降低了高光强对金银花叶片细胞的损伤。处理35 d时,CK组SOD活性略低于Ph 7和Ph 10处理组,分析可能是处理后期自然光照时间(Ph 13)集聚过多能量,间接引发高温胁迫,SOD活性受到抑制。

图1 光周期处理对金银花叶片SOD活性的影响
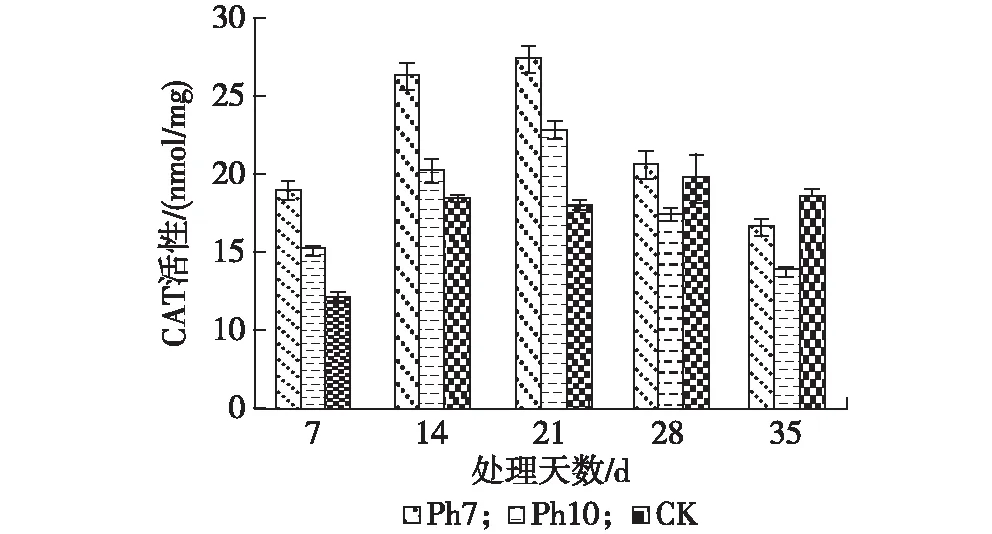
图2 光周期处理对金银花叶片CAT活性的影响
2.3.2 CAT活性 不同光照时间处理对金银花叶片中CAT活性具有一定的影响。表现为短日照处理(Ph 7和Ph 10处理组)CAT活性高于CK组;而处理时间为28—35 d时,Ph 10处理组叶片中CAT活性低于CK组,而Ph 7处理组在处理28 d时略高于CK组,在处理35 d时则低于CK组。
2.3.3 POD活性 由图3可看出,不同光照时间处理下,金银花叶片中POD活性变化趋势与CAT变化相似,表现为Ph 7和Ph 10处理组POD活性高于对照组;在28—35 d时,光照时间10 h处理组POD活性低于Ph 7处理组和对照组。比较同一处理组不同时间POD活性变化趋势发现,2个处理组和对照组POD活性随处理时间延长而呈先升高后降低的趋势,但达到峰值时间不同。
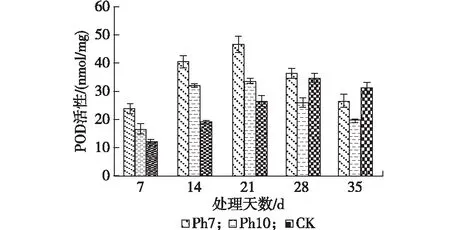
图3 光周期处理对金银花叶片POD活性的影响
3 结论与讨论
3.1 光周期对金银花叶片光合特性的影响
叶片是植物吸收光能进行光合作用的场所,光周期的变化直接影响其光合色素合成及光合速率的高低,从而影响植物生长发育[12]。张欢等[13]研究表明油葵芽苗菜叶绿素和类胡萝卜素含量随着光周期从0到16 h/d的延长显著提高。本研究结果与之相符,即短日照条件下金银花叶片中Chla,Chlb,Chl(a+b)和Car含量均呈降低趋势。推测可能是由于参与叶绿素和类胡萝卜素等光合色素合成的多个酶均属于光响应酶[14],短日照处理组金银花叶片接受光照时间较短,抑制了此类光响应酶的催化活性,进而影响光合色素在叶片中的合成。
光合色素含量直接影响植物光合作用的强弱。李冬梅[15]研究表明,短日照处理可引起桃叶片Pn,Gs降低,Ci升高,认为短日照处理可能导致植株体内积累过多的活性氧,进而破坏光合器官结构和功能,引起叶片衰老,促使叶片维持较低Gs、较高Ci状态。本研究则发现金银花叶片Pn,Gs,Ci及Tr随着光照时间减少而降低,Ls呈升高趋势。分析原因可能是,(1)气孔导度下降引起CO2供应量减少,最终导致叶片光合速率降低,光合产物合成受阻;(2)短日照处理影响了叶片光合色素合成,进而引起光合速率降低。
3.2 光周期对金银花叶片抗氧化酶活性的影响
抗氧化酶又称为保护酶,主要包括SOD,CAT,POD,是植物适应逆境的重要生理指标,其活性的高低是衡量植物适应逆境能力的强弱。周天华[16]研究光照时间对桃树叶片抗氧化系统的影响,发现长日照不同程度降低了SOD,CAT和POD活性,而短日照则激发了3者的活性,认为短日照处理可导致叶片细胞膜脂过氧化损伤。本研究发现,处理初期短日照不同程度地提高了SOD,CAT和POD活性,推测可能是由于短日照对于金银花生长具有胁迫作用,抗氧化酶活性的提高可有效清除植物体内的超氧自由基,增强金银花抗胁迫能力。而随着光照时间延长,植株进行光合作用时间随之延长,可促进植物产生较多的光合同化物,延缓叶片衰老,使得活性氧的积累和清除趋于动态平衡状态。
处理后期(28—35 d),短日照处理金银花叶片中,3种酶活性与对照处理相比增加不显著,特别是光照处理10 h,3种抗氧化酶活性维持较低水平。推测可能是由于随着处理后期南京地区日光照强度增加,减少日照时间可有效减轻强光对叶片的损伤,减少超氧自由基的产生,从而降低3种酶的活性;而长日照处理则因日光照量增加而引发高温、强光胁迫,使得叶片细胞产生过量超氧自由基,导致叶片抗氧化酶活性受到抑制。
