重金属铜对植物毒害机理的研究现状及展望
2018-09-10公勤康群王玲李兆华
公勤 康群 王玲 李兆华
摘要:铜是植物正常生长发育所必需的微量营养元素,随着铜用途的扩大及用量的增加,含铜污染物的排放量逐渐增多,导致植物中铜含量超标,已严重影响到人类健康和生态系统稳定。文章综述了国内外关于铜对植物生長、细胞膜系统、抗氧化物酶系统、光合作用、矿质营养吸收和运输等生理响应等方面的研究进展,指出目前关于铜对植物毒害的研究主要集中在植物的耐铜阈值、毒害生理机理和植物对铜的解毒效应等方面,并提出今后应从拓宽植物研究种类、铜胁迫差异蛋白和基因表达鉴定及维持细胞内稳态的重金属转运体的鉴定等方面进行深入研究,为揭示铜对植物的毒害机理提供依据。
关键词:重金属铜;毒害机理;研究现状;展望
中图分类号:S153.61;Q945.78 文献标志码:A 文章编号:2095-1191(2018)03-0469-07
0引言
铜(Cu)是一种植物正常生长发育所必需的微量营养元素,也被世界卫生组织列为人类和动物所必需的14种微量元素之一。随着铜的用途扩大及用量增加,含铜污染物排放越来越多,根据国家《污水综合排放标准》(GB 8978-1996)规定,排入《地表水环境质量标准》(GB 3838-2002)Ⅲ类水域的污水执行《污水综合排放标准》(2015)一级排放标准,以铜≤0.5 mg/L为达标。2014年《全国土壤污染调查公报》显示全国土壤总的超标率为16.1%,其中铜的点位超标率为2.1%,位居第四。另据报道,我国多地存在铜污染现象,对生态环境及居民健康均构成较大的潜在危害(姚黎霞等,2013;李雪峰,2014;袁文淼,2015;郭爱珍等,2016;朱旭彬等,2016)。铜污染已成为农业生产中急待解决的问题之一。文章对国内外有关铜对植物生长生理毒理的研究进展及研究前景进行归纳,以期为今后开展铜对植物毒害机理的深入研究提供借鉴。
1土壤铜的来源及其毒害机理
1.1土壤铜的来源
土壤铜的来源非常广泛,可分为自然来源和人为输入两种。在自然来源中,成土母质和成土过程对土壤铜含量影响较大(郑喜坤等,2002);人为输入中,工业、交通、农业生产等是引起土壤铜含量超标的重要原因(sfinchez-Pardo et a1.,2012)。人为输入来源主要分为五个方面:一是大气沉降,工业生产、汽车尾气排放及汽车轮胎磨损等产生大量含铜有害粉尘,通过自然沉降和雨淋沉降进入土壤;二是高铜含量饲料添加剂、含铜杀菌剂(蓝矾、波尔多液等)的施用,明显增加了农田中的铜含量;三是农田灌溉中大量使用工业废水、城市污水,许多铜离子(Cu2+)进入土壤(郑喜坤等,2002);四是含铜量较高的污泥被作为有机肥料及土壤改良剂用于农业生产(刘敬勇等,2010);五是畜禽粪便在农业生产中的施用,进入畜禽体内90%的铜会随粪便排出,施入土壤中造成铜在土壤中累积。由于铜具有隐蔽性强、残留时间长、不易降解、毒性强、不可逆的特点,能通过直接接触或食物链传递在生物体内富集,因此已成为国内外瞩目的热点环境问题(关天霞等,2011)。
1.2铜对植物体的毒害机理
铜是某些金属蛋白酶的结构元素,参与植物重要的生物过程,如光合作用、呼吸、氧化超氧化物清除、乙烯传感、细胞壁的新陈代谢和木质化等。但cu2+过量后表现为高毒性,会对植物体产生氧化胁迫,形成有害的活性氧(ROS),破坏细胞膜的结构和功能,同时催化苔腾(Fenton)反应生成羟基自由基,导致脂类、蛋白质和DNA损害(Drazkiewicz et a1.,2004),导致植物出现生物量减少、根系生长受抑制及萎黄病、烫伤、坏死等症状(chaoui and Jarrar,2004)。铜离子的毒性也可导致铁吸收减少(Cha-oui et a1.,2004),干扰植物代谢,如叶绿体完整性丧失、质体膜成分改变及对光合电子传递的抑制(Quartacci et a1.,2000;Pfitsikkfi et a1.,2002)。但在拟南芥(Arabidopsis thaliana)中,上调的大多数基因并不是特定于铜金属的基因,此类基因也被许多其他压力所诱导,如暴露于臭氧、盐、冷和渗透休克(Weber et a1.,2006;Zhao et a1.,2009)。这可能是由于Cu2+引发形成大量ROS,对大多数生物和非生物胁迫的共同反应(zhao et a1.,2009);也可能是为了防止Cu2+诱导的损伤,植物建立了复杂的机制,以避免细胞中游离Cu2+的积累。
2铜对植物生长生理的影响
2.1铜对植物生长的影响
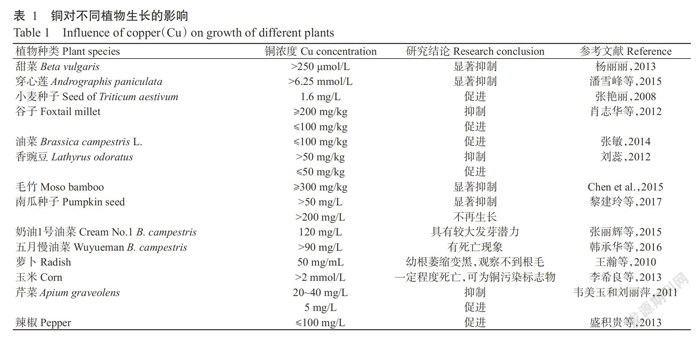
铜是某些金属蛋白酶的结构元素,但过量的铜会导致植物生物量减少、生长发育迟缓,甚至引起萎黄病和坏死。目前,国内外学者研究了多种植物在铜污染条件下的耐性阈值,结果(表1)表明,不同植物对铜的耐受性存在差异,低浓度的铜能够促进植物种子萌发和生长,但当铜浓度继续增加后,对植物生长产生不同程度的抑制作用。
2.2铜对植物细胞膜系统的影响
细胞是植物与外界环境物质交换、信息交流的屏障。重金属浓度大于植物生长的阈值时,会导致植物发生毒害效应,致使细胞膜结构和功能被破坏(Mahalingam and Fedroff,2003)。研究表明,Cu2+浓度为100 mg/kg时已超出萝卜幼苗根系对铜的耐受限度,其相对电导率、丙二醛(MDA)含量增加,细胞膜系统受损(韩春梅,2010);Cu2+浓度为160 gmol/L时,费菜的相对电导率和MDA含量最小,Cu2+浓度为400 gmol/L时,其相对电导率及MDA、脯氨酸和可溶性糖含量均与对照差异显著(袁红艳,2010);Cu2+浓度<.200 mg/kg可促进香椿的生长,随着Cu2+浓度的升高,MDA含量先降低后升高,可溶性蛋白含量先降低后升高而后又降低,可溶性糖含量先降低后升高(李晓晶等,2013);蓖麻叶片中的MDA含量和细胞膜透性在铜<.200 gmol/L时无显著变化,高浓度铜处理时二者显著升高(李春雷,2016)。
2.3铜对植物抗氧化物酶系统的影响
在胁迫条件下时,植物体ROS的产生和清除平衡遭到破坏,加速了ROS的积累,进而影响植物正常的生长发育。植物通过调节体内的抗氧化物酶系抵抗Cu2+脅迫,当Cu2+浓度超过植物耐铜阈值时,其抗氧化物酶系被破坏。研究发现,Cu2+胁迫会导致人参根部的葡萄糖6-磷酸脱氢酶(G-6-PDH)、莽草酸酯脱氢酶(SKDH)、苯丙氨酸氨裂解酶(PAL)和肉桂醇脱氢酶(CAD)活性增加,酚醛和木质素积累(Aliet a1.,2006);在10umol/L的CuSO4胁迫下,“常州乌塌菜”不结球白菜体内的维生素(ASA)含量、过氧化氢酶(CAT)和过氧化物酶(POD)活性均高于“二青”不结球白菜,表现出较强的耐铜性(宋玉萍,2008);Cu2+浓度为30 mg/L时,小麦幼苗中的超氧化物歧化酶(SOD)活性最高,是对照的3.67倍(李春喜等,2011);速生柳苗可通过增加保护酶活性和脯氨酸含量来提高其对Cu2+的耐受力(刘治昆等,2011);Cu2+浓度为50和70 mg/L时,小麦种子通过增加CAT和SOD活性缓解铜胁迫,但Cu2+浓度为100和150 mg/L时,小麦种子中的POD、CAT和SOD活性及MDA含量均明显升高,阻碍其生长发育(薛盈文等,2016);在铜胁迫下,小麦体内的超氧自由基(O-2)含量、POD活性均呈先升高后下降的变化趋势,SOD活性呈升高趋势(赵庆芳等,2017)。
2.4铜对植物光合作用机理的影响
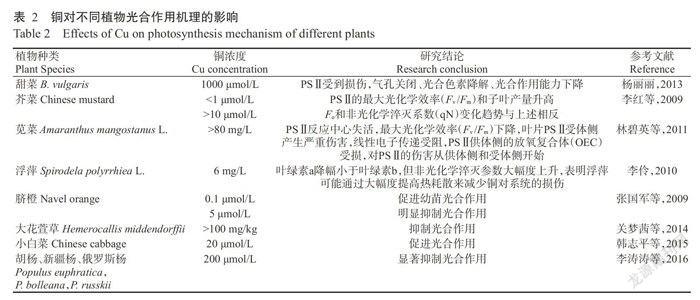
重金属对植物光合作用的抑制机理一般是降低叶绿素含量,损伤叶绿体结构和膜系统,导致叶片光合酶损伤,抑制光合和非光合磷酸化、RUBP羧化酶活性和光系统的电子传递(Baryla et a1.,2001)。已有研究(表2)表明,铜作为植物生命活动的微量元素,当其浓度较低时能够促进光合作用,反之则抑制。过量的铜会改变叶绿体或类囊体的结构,甚至会导致植物生物量减小,细瘦萎黄,铜和D1蛋白的络氨酸残基相结合,可抑制光系统II(PSⅡ)的电子传递(Burda et a1.,2002),或导致细胞色素b559氧化还原端的改变(Arellano et a1.,1995),光合作用所储存的能量减少,光合电子传递降低,导致生物量减少(Burda et a1.,2003),铜胁迫使叶绿素含量大幅下降,24 h内使初始荧光(Fo)和最大荧光(Fm)升高,特别是Fm可能出现负值,表明光合器官己受到破坏(李红敬等,2003)。
2.5铜对植物矿质营养元素吸收和运输的影响
通过测定植物中重金属的浓度和富集系数两个指标可了解重金属在植物体内的吸收和积累情况,从而判别植物受重金属污染所产生的毒性效应。在矿质营养的吸收方面,研究发现,除钠和锌外,铜对小麦幼苗中钾、钙、镁、铁、锰、硼六种矿质元素的吸收存在明显拮抗作用(张志娟,2011);硒能促进小白菜根系中的铜向地上部转运,高含量铜胁迫使更多的硒滞留在小白菜地下部分(胡斌等,2011)。除了对矿质营养吸收存在影响外,铜在植物体内富集的部位多在细胞壁。在铜胁迫下,小麦幼根细胞壁是铜的主要储存库,其次为细胞质(张志娟,2011);豆苗叶片细胞壁是cu2+积累的主要部位(Bouazizi eta1.,2011)。海州香薷根细胞壁的羟基、羧基和氨基是Cu2+的主要结合位点,细胞壁中果胶和纤维素是铜离子主要吸附结合部位,分别吸附了19.85%和25.48%的铜,细胞壁多糖在海州香薷对铜解毒方面发挥着重要作用(刘婷婷,2014)。
3展望
铜广泛存在于自然界,是植物生长、工业生产、农药合成、饲料加工等方面必不可少的元素之一,也是参与植物多种生命物质合成的重要元素。但由于生产中过度追求高产量、高品质、低成本、易管理的商品标准和要求,过量使用农药、化肥、饲料添加剂,超规排污造成土壤、水资源中铜含量超标,严重影响到人类健康和生态平衡。目前,重金属对植物的毒害机理是国内外学者研究的热点,有关铜对植物的毒害机理研究现状及研究趋势主要集中在以下3个方面:
(1)植物的耐铜阈值及筛选具有较强铜富集效应的植物。前人已对芥菜(Chigbo et a1.,2013)、海州香薷(刘婷婷,2014)、向日葵(Bortolon,2015)、毛竹(Chen et a1.,2015)和金盏花(Goswami and Das,2016)等植物的铜富集效应进行研究,得出铜胁迫下植物正常生长的最适浓度,以及利用铜富集效应修复铜污染土壤,但针对耐铜性植物及具有富集效应植物的研究范围过于狭窄,主要集中在20-30种常见植物,需要加强对其他植物尤其是直接进入人类食物链中植物的研究,以便更好修复和利用铜污染土壤。
(2)铜对植物毒害的生理机理。铜胁迫对小麦(张艳丽,2008;李春喜等,2011;Sfinchez-Pardo eta1.,2012;赵庆芳等,2017)、、辣椒(盛积贵等,2013)油菜(张敏,2014;张丽辉等,2015)、小白菜(韩志平等,2015)和南瓜(黎建玲等,2017)等植物毒害的生理机理已有较多研究,包括植物生长特性、细胞膜系统、抗氧化物酶系统、光合作用机理、矿质元素吸收和运输等宏观水平,以及细胞亚显微结构、蛋白质合成、核酸代谢和基因表达等微观水平。今后,还可进一步加强研究Cu2+在植物亚细胞水平上的迁移、转运、积累及铜胁迫差异蛋白和基因表达的鉴定等,以更全面、系统地诠释铜对植物的毒害机理。
(3)植物对铜毒害的解毒效应。已有研究表明,小白菜(胡斌等,2011)、玉米(李希良等,2013)、牡丹(Jin et a1.,2015;Staehlin et a1.,2016)、牛蒡(Jung et a1.2015)等植物可通过液泡聚集Cu2+、调控铜基因表达、提供Cu2+结合位点等机理缓解铜毒害。今后还应从维持细胞内稳态的重金属转运体的鉴定及耐铜基因的鉴定、提取和转移等方面深入研究植物对铜毒害的解毒效应。
(责任编辑 王晖)
