腐皮镰孢霉菌侵染及保鲜剂处理对秋葵相关抗性酶的影响
2018-09-10辛松林秦文孙传红徐丹黄倩毛美丹方子豪
辛松林 秦文 孙传红 徐丹 黄倩 毛美丹 方子豪
摘要:为研究秋葵果实响应外界胁迫反应及合理利用果实抗性酶的保护功能,以川秋葵为试验材料,采用人工接种腐皮链孢霉菌和保鲜剂处理的方法,研究秋葵果实抗性酶的变化。结果表明,POD、PPO、PAL、C4H、4CL、CAD和TAL参与了秋葵果实响应腐皮链孢霉菌侵染和成熟衰老的反应过程。4CL、POD、PPO在整个腐皮链孢霉菌侵染阶段活性持续增高起着重要的防御作用;PAL、CAD在腐皮链孢霉菌侵染的4d内活性增高,且在整个侵染阶段都保持了较高的活性:C4H在腐皮链孢霉菌侵染的3d内起着重要的防御作用:TA,在侵染24 h迅速启动响应反应。不同保鲜剂对抗性酶活性表达的诱导作用不同,1-MCP与壳聚糖复合处理对维持果实抗病性和采后品质的效果最好。
关键词:川秋葵;腐皮链孢霉菌;保鲜剂;抗性酶
中图分类号:S649
文献标识码:A
文章编号: 1000-4440(2018)05-1161-08
秋葵,又称黄秋葵,属于锦葵科秋葵属一年生草本植物,由于其富含果胶、黄酮及人体必需氨基酸,具有一定的保健功能,因此广受消费者喜爱,然而由于秋葵表面积大,果皮表面附有细密绒毛,很容易受到病原菌的侵染而加速秋葵的腐败变质,影响了贮藏时间。秋葵受到病原菌侵染后,组织会产生一系列复杂的生理生化变化,以维持自身代谢的动态平衡,增强对病原菌的抵抗能力。其主要机理为:病原菌通过侵染健康组织,诱导产生活性氧分子,大量积累的活性氧对植株造成氧化伤害。另一方面,植株自身的抗氧化防御系统同时发生作用,植株体内的抗氧化物酶类如过氧化物酶(PPO)、多酚氧化(POD)等通过清除活性氧分子,从而诱导植株产生适应性响应。此外,苯丙烷代谢系统作为生成酚类物质的主要次生代谢途径,在植株抗逆境胁迫和适应性防御反应中起着重要作用。
秋葵果实对不同病原菌会表现出不同的抗感性,课题组前期对诱导秋葵果实在贮藏过程中腐败的病原菌进行了研究,结果表明,腐皮链孢霉菌侵染是诱导果实腐败的主要原因,并分离纯化出腐皮链孢霉菌。腐皮镰孢霉菌是常见的土壤习居菌,春天气温回升,土壤湿润,菌核开始萌发产生子囊盘和子囊孢子,从而侵染植株根茎部或基部叶片及其他组织,发病后产生菌丝,受害病叶与邻近健株接触即可传病。本试验研究腐皮镰孢霉菌侵染前后及保鲜剂处理对秋葵相关抗性酶的影响,探讨秋葵在致病菌侵染和保鲜过程中相关抗性酶的变化规律,为进一步研究秋葵果实响应外界胁迫反应及合理利用果实抗性酶的保护功能提供基础。
1 材料与方法
1.1 材料与试剂
秋葵(品种为川秋葵):四川省植物工程研究院提供。选择无伤口、无病虫害、果荚长度6.0~8.0cm的果实,采摘后及时置于2~4℃保鲜库中预冷备用。
腐皮镰孢霉菌孢子悬浮液配制:腐皮镰孢霉菌从贮藏过程中发病的秋葵果实中分离获得,经纯化培养后,在PDA培养基上28℃恒温培养6d,转入含有10ml 0.01% Tween 20无菌水的50ml三角瓶中,在微型旋涡混合器上振荡15s,再用双层纱布过滤,滤液用血球计数板计数,算出孢子悬浮液的含菌量后,最终稀释至含菌量为1ml lxl05孢子的孢子悬浮液,备用。
主要试剂:壳聚糖(脱乙酰度≥90%),购于成都科龙化工试剂厂:安喜布(有效质量浓度0.45mg/L,规格为25cmx20cm),购于兰州嘉诚生物技术有限公司:其他试剂均为市售分析纯。
1.2 仪器与设备
UV-3200扫描型紫外/ 可见分光光度计(上海美谱达仪器有限公司产品),冷冻高速离心机(美国Thermo公司产品),恒温水浴锅,电子天平,培养箱等。
1.3 试验方法
1.3.1 腐皮链孢霉菌侵染处理 选取预冷后的果实,将秋葵用75%酒精进行消毒,再用无菌水进行清洗,沥干水分待用。用灭菌过的打孔器(直径为1.0cm)在秋葵赤道部位表面打8个深度为1.0~1.5cm的孔,其中5个孔不接菌,其余5个孔中分别接入10μl含菌量为lml lxl05孢子的腐皮镰孢霉菌孢子悬浮液,再用保鲜膜包住打孔部位人泡沫包装箱,室温(250C+2℃,湿度85%~ 95%)条件下贮藏,分别于接菌1d、2d、3d、4d、5d取樣,进行相关指标的测定。
1.3.2 保鲜样品处理 将挑选好的秋葵分为4组,每组250个,分别进行以下处理:(1)壳聚糖涂膜处理:将秋葵浸于1.0%的壳聚糖涂膜液中60s,待秋葵果实表面完全浸润,捞出后自然风干;(2)l—MCP处理:将0.5片安喜布和250个秋葵同时放人带有纸屑的泡沫箱(340mmx220mmx180mm)中,盖紧盖子,放置24h后取出秋葵;(3)1-MCP与壳聚糖复合处理:按照(1)的方法先处理秋葵,然后按照(2)的方法再次处理秋葵;(4)对照:以不做任何处理的秋葵为对照。
将上述3个处理和1个对照的秋葵置于常温(20℃+1℃)条件下贮藏,相对湿度为85%~ 90%,每24 h取样进行各相关指标的测定。
1.3.3 指标测定 苯丙氨酸解氨酶(PAL)活性测定参照Liu等的方法,酶活性单位为U/(h·g),FW。4一香豆酰一辅酶A连接酶(4CL)活性测定参照朱明华等的方法,酶活性单位为U/(min·g),FW。肉桂酸一4羟化酶(C4H)活性测定参照Lamb和Rubery的方法,酶活性单位为U/(min·g),FW。肉桂醇脱氧酶(CAD)活性测定参照Momson等的方法,酶活性单位为U/(h·mg),FW。酪氨酸解氨酶(TAL)活性测定依据Wajahatullah Khan等的方法,酶活性单位为U/(h·mg),FW。过氧化物酶(POD)活性测定参照Jiang等的方法,酶活性单位为U/(min·g),FW。多酚氧化酶(PPO)活性测定参照Clairbone等的方法,酶活性单位为U/(min·g),FW。
1.4 数据处理
每组处理进行5个平行实验。试验数据采用SPSS20.0软件和Origin 8.1软件进行分析处理,选用ANOVA进行邓肯氏多重差异显著性分析。
2 结果与分析
2.1 腐皮链孢霉菌侵染及保鲜剂处理后秋葵果实POD活性的变化
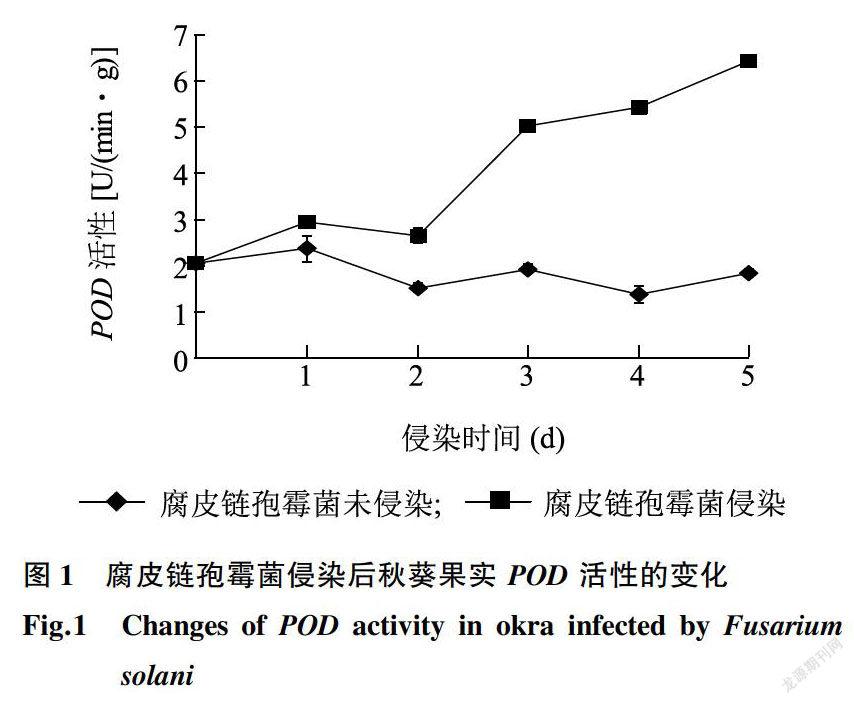
由图l可知,在贮藏期内,腐皮链孢霉菌未侵染组POD活性变化缓慢,变化幅度较小;腐皮链孢霉菌侵染组POD活性呈持续上升的趋势,POD活性在侵染第5d达到最大值,为6.43 U/(nun·g),FW,极显著高于未侵染组(P<0.01)。这种现象的原因可能是秋葵受到腐皮链孢霉菌侵染后,迅速启动相关防御体系,果实POD活性上升以保持秋葵体内活性氧代谢平衡,维持膜结构完整性,抵御病原菌对秋葵果实的侵害。
由图2可知,贮藏期间,空白组的POD活性变化幅度较小.3个保鲜剂处理组的POD活性呈先升高后下降的变化趋势,在贮藏第ld,1-MCP处理组和复合处理组的POD活性达到最大值,复合处理组POD活性与对照组差异达极显著水平(P<0.01),1一MCP处理组POD活性与对照组差异显著(P<0.05):在贮藏第2d,壳聚糖处理组的POD活性达到最大值,与对照组差异显著(P<0.05),说明1-MCP和壳聚糖能够诱导POD的活性提高。随着贮藏时间的延长,保鲜剂处理组的POD活性下降,在贮藏第Sd,对照组的POD活性高于1-MCP处理组和壳聚糖处理组。可能是因为贮藏前期保鲜剂处理能够有效提高POD活性,及时清除细胞内部的自由基,壳聚糖本身也是一种广谱杀菌剂,这对提高秋葵适应性防御能力非常重要:贮藏后期,由于病原菌的侵染和组织老化,对照组上调POD活性以响应组织氧化和病原菌侵染的反应过程。
2.2 腐皮链孢霉菌侵染及保鲜剂处理后秋葵果实PAL活性的变化
腐皮链孢霉菌侵染诱导PAL活性增强。由图3可知,PAL在腐皮链孢霉菌侵染的第ld迅速启动响应反应,活性迅速升高,随后3d活性缓慢上升,在第4d达到最大值,为638.622 U/(h·g),FW,随后下降;未侵染组PAL活性呈先升后降趋势,在第4d出现最大值为454.895 U/(h·g),FW。PAL是苯丙氨酸代谢途径的限速酶,参与了多种激发子诱导的抗性,其活性增强是一种自我保护机制的反应,有助于抵御病原菌的侵染。
由图4可知,PAL活性呈现波动性变化。贮藏前2d,3个保鲜处理组的PAL活性高于对照组。贮藏第3d,壳聚糖处理组的PAL活性低于对照组,并在此后保持相对较低的水平。这可能与壳聚糖对腐皮链孢霉菌孢子萌发和菌丝体生长具有抑制作用有关。贮藏第4d,1-MCP处理组PAL活性低于复合处理组和对照组,这可能与1-MCP作为植物生长调节剂诱导PAL表达有关。复合处理组的PAL活性在第4d达到最大值532.5U/(h·g),FW,这单纯从1-MCP处理或是壳聚糖处理的结果很难解释,推测有可能与二者协同作用有关,诱导因子促进酶的合成是由于刺激了酶蛋白mRNA的形成。总体而言,在贮藏初期,保鲜剂处理对于PAL的活性有显著提高作用,可以提高秋葵内部植保素和木质素的生成速率,抵御组织的老化以及病原菌胁迫,随着贮藏时间延长,秋葵组织内部苯丙烷代谢趋于平衡稳定状态,保鲜剂对PAL活性的影响不大。
2.3 腐皮链孢霉菌侵染及保鲜剂处理后秋葵果实PPO活性的变化
由图5可知,腐皮链孢霉菌未侵染组PPO活性在侵染期间呈先降后升的趋势,侵染第3d的PPO活性最低,侵染第Sd的活性最高,为0.5489U/(min·g),FW。其变化原因可能是随着贮藏时间的延长,秋葵组织衰老,产生活性氧的积累,诱导PPO活性升高去除自由基,延缓果实衰老进程。腐皮链孢霉菌侵染组果实PPO活性呈现连续上升的趋势,在侵染第5d达到最大值,为0.6465U/(min·g),FW,与腐皮链孢霉菌未侵染组差异不显著(P>0.05)。结果表明,PPO参与了秋葵果实响应腐皮链孢霉菌侵染的反应过程,在整个侵染期间起着重要的防御作用。
由图6可知,1-MCP处理组和复合处理组在贮藏期间的PPO活性变化呈持续上升趋势,在第Sd达到最大值;壳聚糖处理组在贮藏期间的PPO活性呈现先升后降的趋势,在第3d达到最大值。在贮藏第3d,保鲜处理组PPO活性均极显著高于对照组(P<0.01),可能是因为保鲜剂能誘导秋葵组织内部存在的PPO活性迅速上升,促进木质素以及抗菌醌类物质的形成,对植物组织起到一定的防御保护作用。由此可见,保鲜处理可以有效提高PPO活性。在贮藏后期,壳聚糖处理组PPO活性下降,这种现象的原因可能是随着贮藏时间延长,调控PPO各个基因表达不同,活性随之变化。对照组PPO活性在贮藏前3d变化不大,从贮藏第3d开始,PPO活性迅速上升,在第Sd达到最大值,可能由于病原菌的侵染、组织褐变,对照组PPO活性被诱导增强。
2.4 腐皮链孢霉菌侵染及保鲜剂处理后秋葵果实4CL活性的变化
由图7可知,腐皮链孢霉菌未侵染组在整个试验期间4CL活性上升缓慢,活性最大值为34.49U/(min·g),FW;腐皮链孢霉菌侵染组在侵染第1d 4CL活性显著上升,随后下降,第4d秋葵果实4CL活性达到最大值为83.35U/( min·g),FW,与未侵染组差异达极显著水平(P<0.01)。结果表明,4CL参与了秋葵果实响应腐皮链孢霉菌侵染的反应过程。
由图8可知,随着贮藏时间的延长,对照组和保鲜处理组的4CL活性总体呈上升的趋势,经保鲜剂处理的秋葵果实4CL活性始终高于对照组。复合处理组4CL活性最大值为61.57 U/(min·g),FW,1-MCP处理组4CL活性最大值为45.92 U/(min·g),FW,壳聚糖处理组4CL活性最大值为46. 27U/(min·g),FW,对照组4CL活性最大值为34.49U/(min·g),FW,复合处理组与对照组的4CL活性差异达极显著水平(P<0.01),1-MCP处理组、壳聚糖处理组与对照组的4CL活性差异不显著(P>0.05)。
感官评价分别从色泽、质地、外观和接受度等方面进行评价。由图9可知,经保鲜剂处理的秋葵果实经过S d的贮藏,其品质显著优于对照组。可能因为4CL控制着苯丙烷主途径向分支途径的转折,其代谢产物——酚类物质可氧化成能对病原菌产生直接毒性的醌类物质,随着酚类物质和木质素的积累,能有效抑制病原菌的扩展。
2.5 腐皮链孢霉菌侵染及保鲜剂处理后秋葵果实C4H活性的变化
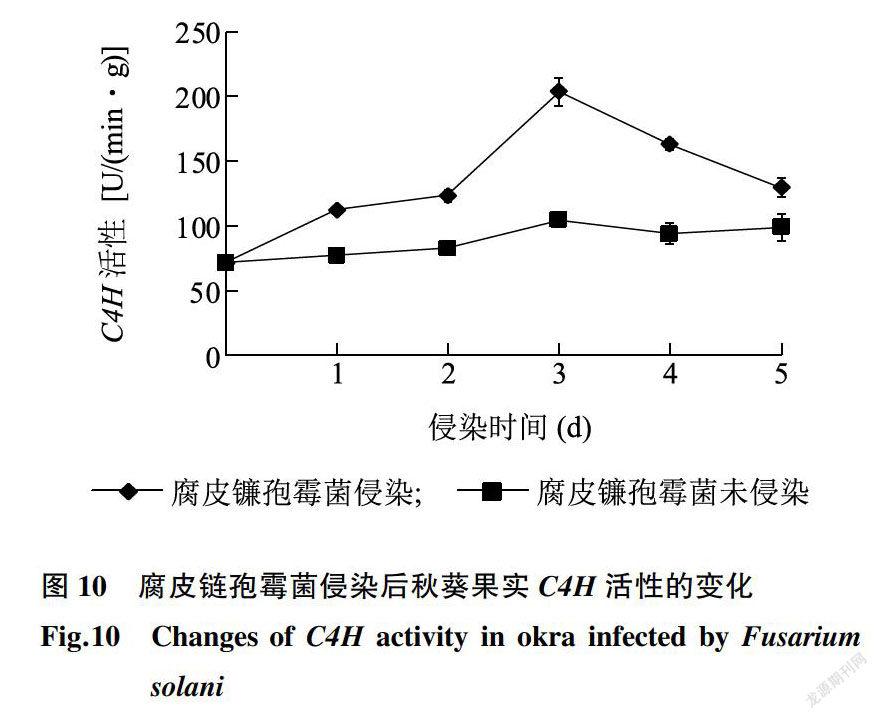
C4H是苯丙烷代谢的关键酶。由图10可知,腐皮链孢霉菌侵染秋葵果实后,C4H活性显著上升,在侵染第3d达到最大值,为203.74U/(min·g),FW,随后下降。未侵染组C4H活性在整个贮藏期间的上升趋势缓慢,在贮藏第Sd达到最大值,为98.56 U/(min·g),FW。
与腐皮链孢霉菌未侵染组相比,侵染组C4H活性上升明显,这种现象可能的原因是秋葵受到腐皮链孢霉菌侵染后,果实产生系统抗性,诱导C4H活性增强,有利于苯丙烷代谢酚酸类物质的合成,由此形成的香豆酸、阿魏酸和咖啡酸等酚酸,抵御腐皮链孢霉菌的侵染。
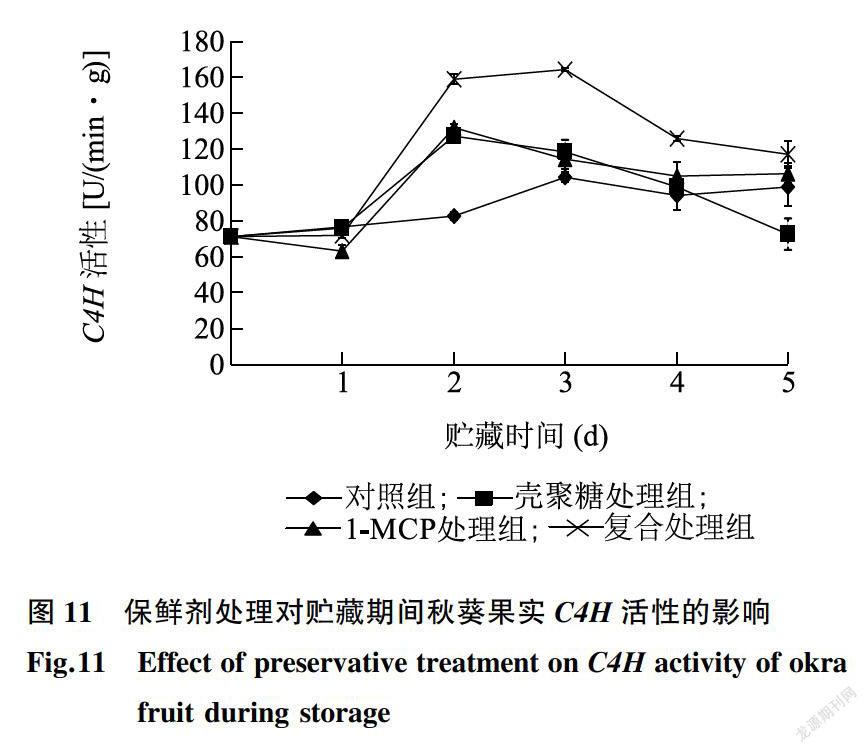
由图11可知,对照组的C4H活性在贮藏前期呈波动式上升趋势,在贮藏第3d,C4H活性出现最大值,为104.1U/(min·g),FW。保鲜剂处理组呈现先升后降的趋势,壳聚糖处理组和1-MCP处理组均在贮藏第2d,C4H活性达到最大值,分别为127.0U/(min·g),FW和132.0U/(min·g),FW,而复合处理组则在贮藏第3d时C4H活性达到最大值,为164.0U/(min·g),FW。结果表明,C4H参与了秋葵果实响应腐皮链孢霉菌侵染和保鲜剂处理的反应过程,C4H活性升高是秋葵果实抵抗腐皮链孢霉菌侵染和延长贮藏期的重要机制之一。
2.6 腐皮链孢霉菌侵染及保鲜剂处理后秋葵果实CAD活性的变化
由图12可知,腐皮链孢霉菌侵染组的CAD活性呈现先升后降的趋势,第4d急剧上升,CAD活性达到最大值,随后CAD活性下降;而未侵染组呈逐步上升趋势,在贮藏后期的CAD活性上升较快,可能是因为秋葵果实的老化诱导CAD活性上调。在整个侵染期间,侵染组的CAD活性较未侵染组高,说明腐皮链孢霉菌侵染能够诱导秋葵果实上调CAD活性以抵抗侵染。
由圖13可知,对照组的CAD活性呈上升趋势,在贮藏第3d急剧上升,活性最大值为338.28U/(h·mg),可能因为秋葵果实的快速衰老诱导了CAD活性的急剧上升。壳聚糖处理组和复合处理组的变化趋势基本一致,呈逐步上升趋势,最大值分别为135.98 U/(h·mg)和328.86 U/(h·mg)。1一MCP处理组的CAD活性呈现先升后降的趋势.CAD活性第3d达到最大值,为131.82 U/(h·mg)。结果表明,在贮藏前期,保鲜剂均可诱导秋葵果实CAD活性升高,提高抗病性,延缓果实衰老;贮藏中后期,由于果实的衰老,病原菌侵染,诱导了对照组CAD活性表达更高。
2.7 腐皮链孢霉菌侵染及保鲜剂处理后秋葵果实TAL活性的变化
由图14可知,腐皮链孢霉菌侵染组的TAL活性呈波动式上升下降趋势,未侵染组TAL活性呈现先升后降的变化趋势,侵染组TAL最高活性与未侵染组TAL最高活性差异达极显著水平(P<0.01)。TAL与PAL都属于芳香族解氨酶家族,TAL能不经C4H进行非氧化脱氨,直接将L-酪氨酸(L-Tyr)转化为香豆酸,转化过程中的某些变化可能使得TAL活性出现波动。
由图15可知,复合处理组和1-MCP处理组TAL活性在贮藏前4d呈持续上升趋势,复合处理组TAL活性最大值达到65.11 U/(h·mg),1-MCP处理组最大值为58.03U/(h·mg);壳聚糖处理组TAL活性呈波动式上升,最大值出现在贮藏第4d,为42.09U/(h·mg);对照组TAL活性呈先升高后降低的趋势,最大值出现在贮藏第2d,为19.52U/(h·mg)。
3 讨论
果实组织采收后由于病原菌侵染会产生一系列复杂的生理生化变化,以维持自身代谢的动态平衡,诱导抗病性是采后果实贮藏保鲜的重要机制之一。果蔬中含有大量的抗性酶,在成熟衰老、病原侵染、机械损伤过程中都能诱导抗性酶做出有效响应,但是不同环境下抗性酶的变化规律是不一样的。4CL、POD、PPO在整个腐皮链孢霉菌侵染阶段活性持续增高起着重要的防御作用;PAL是苯丙烷类代谢途径中与抗菌功能产物形成相关的关键酶和限速酶,为木质素和植保素的合成提供前体,在植物早期防卫反应中具有重要的作用。PAL在腐皮链孢霉菌侵染的4d内活性持续增高,且在整个侵染阶段都保持了较高的活性;在腐皮链孢霉菌侵染试验中,侵染组和对照组的CAD活性在第4d显著上升,且侵染组升高的幅度显著高于对照组。C4H、4CL、CAD和TAL是木质素合成的关键酶,它们活性的高低不仅影响木质素的含量、木质素单体的组成,而且影响酚类物质、植保素、类黄酮等抗菌物质的生物合成,这些次级代谢产物在植物的成熟衰老、抵御病虫害、抗逆反应等方面发挥着重要作用。侵染组TAL活性出现波动性变化可能由于L-酪氨酸(L-Tyr)转化为香豆酸过程中的某些复杂反应所致,有待于对TAL催化机制深入研究。保鲜试验结果表明,抗性酶POD、PPO、PAL、C4H、4CL、CAD和TAL参与了秋葵果实响应成熟衰老和病原菌侵染的反应过程,然而不同保鲜剂对抗性酶活性表达的诱导作用不一样,包括POD、PPO、PAL、C4H、4CL、CAD和TAL的酶活性升高以抵抗秋葵果实的成熟衰老以及病原菌侵染,维持果实的抗病性和采后品质。综合考虑,复合处理组的效果最好。这与马铃薯块茎干腐病、猕猴桃灰葡萄孢霉、杏果实抗病性和厚皮甜瓜采后病害中的研究结果相类似。但是,保鲜剂是如何在分子水平上调控秋葵果实抗性酶表达的机制尚需进一步研究。
本试验通过研究腐皮链孢霉菌侵染黄秋葵后抗性酶POD、PPO、PAL、C4H、4CL、CAD和TAL,活性的变化发现,这些抗性酶在腐皮链孢霉菌侵染的黄秋葵果实中都能进行有效响应,且在未侵染组中不同保鲜剂处理对这些抗性酶活性表达的诱导作用不同,试验结果表明,1.0%的壳聚糖和0.45mg/L的1-MCP复合处理保鲜效果最好。
