4株新出现的Ⅰ类新城疫病毒F基因的变异分析
2018-09-03魏润宇罗瑶瑶王静静郑东霞赵云玲刘文博刘华雷王志亮
魏润宇,罗瑶瑶,吕 艳,王静静,郑东霞,赵云玲,刘文博,刘华雷*,王志亮*
(1.扬州大学兽医学院,扬州225009; 2.中国动物卫生与流行病中心,青岛 266032;3.青岛农业大学,青岛 266109; 4.江苏高校动物重要疫病与人畜共患病防控协同创新中心,扬州 225009)
新城疫(Newcastle disease, ND)是由新城疫病毒(Newcastle disease virus, NDV)强毒株感染引起禽的一种急性、高度接触性烈性传染病,世界动物卫生组织(OIE)将新城疫列为必须报告的动物疫病,我国将其列为一类动物疫病,《国家中长期动物疫病防治规划(2012—2020年)》将其列为优先防控的重大动物疫病。NDV属于副黏病毒科、禽腮腺炎病毒属,是一种有囊膜、单股、负链、不分节段的RNA病毒。新城疫病毒具有遗传学多样性,可分为Class Ⅰ和ClassⅡ两类,每类又可进一步分为多种不同的基因型或基因亚型[1]。Ⅰ类NDV至少包括10种基因型[2],Ⅱ类NDV至少包括18个基因型[3]。Ⅰ类新城疫病毒主要分布于北美野生水禽和亚洲活禽市场中。除1990年在爱尔兰分离到Ⅰ类NDV强毒株外,其余分离株均为弱毒株,但现有研究表明,Ⅰ类NDV弱毒株可通过鸡气囊或鸡胚连续传代后增强毒力[4]。因此,对流行的Ⅰ类NDV进行系统监测、分析病毒的流行规律和变异情况,对新城疫的防控具有重要意义。2008年,我国首次分离到Ⅰ类NDV[5]。近年来主动监测数据表明,Ⅰ类NDV在国内分布广泛,流行毒株以基因3型为主,同时存在少量基因2型和基因1型毒株[6],2015年在广西活禽市场的家鸭中分离到1株新型Ⅰ类NDV[7]。本研究于2016年在我国部分地区活禽市场开展新城疫主动监测,分离到4株新型Ⅰ类NDV,并对分离株进行F基因序列分析和遗传进化分析,为进一步了解我国Ⅰ类NDV的分子特征奠定基础。
1 材料与方法
1.1 样品来源
2016年在全国新城疫主动监测过程中从江苏和宁夏活禽市场中采集的家禽棉拭子样品。
1.2 病毒分离与鉴定
将采集的拭子样品按照常规方法接种9~11日龄SPF鸡胚进行病毒分离与鉴定,将鉴定为血凝阳性的尿囊液与NDV标准血清进行血凝抑制试验,鉴定为NDV阳性的尿囊液于-80 ℃保存。对分离到的NDV采用蚀斑纯化法[8]进行纯化,将纯化后的病毒置-80 ℃保存备用。
1.3 RNA提取、RT-PCR和序列测定
采用Roche的 High Pure Viral RNA Kit提取病毒RNA。参考GenBank中NDV序列,设计了用于鉴定Ⅰ类NDV的特异性引物(表1),利用Primer-ScriptTMOne Step RT-PCR Kit Ver.2(TaKaRa)进行常规RT-PCR扩增,经1.5%琼脂糖凝胶电泳鉴定为阳性的样品,送至大连TaKaRa公司测序。

表1 新城疫病毒鉴定引物及F基因扩增引物
1.4 F基因的RT-PCR扩增
根据GenBank中NDV序列,设计F基因扩增引物序列(表1),利用Super-Script III One-Step RT-PCR PlatinumTaqHiFi(Invitrogen) 扩增F基因,扩增产物送至大连TaKaRa公司测序。
1.5 F基因的序列分析和遗传进化
利用DNAStar软件,对病毒F基因序列进行拼接、编辑和同源性比较。利用MEGA软件,对病毒的F基因与GenBank中公布的Ⅰ类新城疫参考株(表2)进行比对和遗传进化分析。采用邻位相连法(Neighbor-joining)构建系统进化树,bootstrap设置1 000次重复。
表2 CⅠ新城疫病毒信息
2 结 果
2.1 病毒分离与鉴定
样品接种于SPF鸡胚后,经血凝和血凝抑制试验,成功鉴定出4株Ⅰ类新城疫病毒(表3),电泳图见图1,分离株地理分布见表3。
2.2 F基因扩增与序列分析
对4株分离株进行F基因扩增和序列分析,分离株F基因编码区长度均为1 662 nt,共编码553个氨基酸,裂解位点氨基酸序列均为112ERQERL117,具有NDV弱毒株的典型分子特征。4株病毒F蛋白均含有12个半胱氨酸(分别位于aa76、aa199、aa338、aa347、aa362、aa370、aa394、aa399、aa401、aa424、aa514、aa523)和6个糖基化位点(分别位于aa86、aa192、aa367、aa448、aa472、aa542)。4株 分离株与新城疫病毒F蛋白功能区参考序列[9]相比:融合肽(fusion peptide)有5处变异,七肽重复区(HRa、HRb、HRc)各有2处变异,跨膜区(transmembrane domain)有6处氨基酸变异(表4)。通过与50株CⅠ新城疫病毒(表2)F蛋白功能区比较发现:4株分离株与CⅠ的NDV F蛋白功能区氨基酸一致,不存在变异现象(表4)。
同源性分析结果显示,3株宁夏分离株F蛋白氨基酸相似性为100%,与江苏分离株(JS2145)相似性为97.9%。4株分离株与Ⅰ类NDV基因1~10型代表株相似性为80.4%~98.0%,与北美分离株[10](Greater Black-backed Gull/USA/NJ/AI10-835)相似性最高(97.7%~99.3%),而与2015年国内Ⅰ类新型分离株(duck/Guangxi/1261/2015)相似性仅为95.5%~97.0%(表5)。
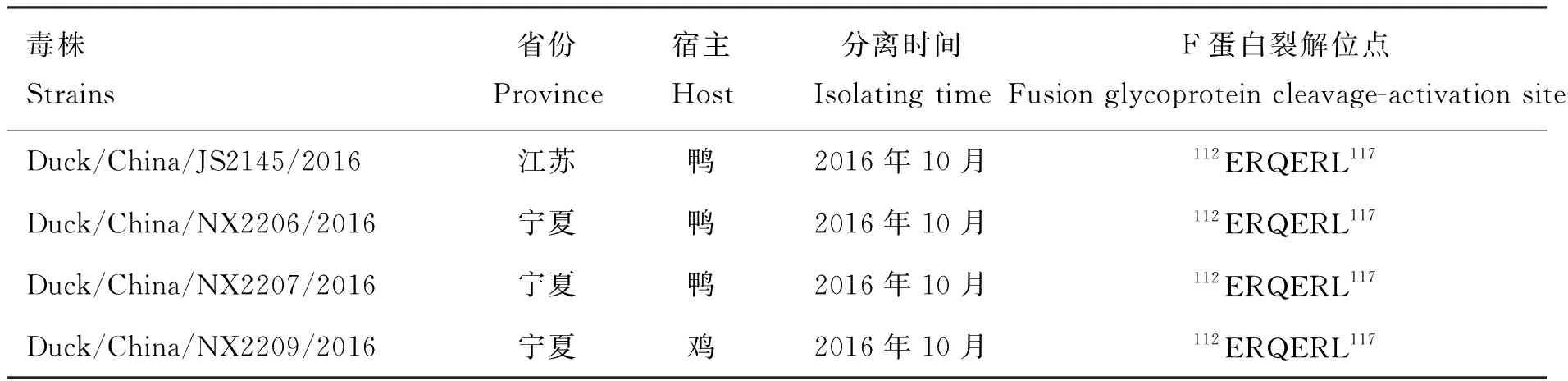
表3 本研究中4株Ⅰ类NDV分离株的详细信息
M. DL2000 相对分子质量标准; 1. JS2145分离株; 2. NX2206分离株; 3. NX2207分离株; 4. NX2209分离株; 5. 阳性样品M. DL2000 marker; 1. JS2145 isolate; 2. NX2206 isolate; 3. NX2207 isolate; 4. NX2209 isolate; 5. Positive sample图1 4株新城疫病毒F基因的RT-PCR扩增Fig.1 RT-PCR amplification of F gene of 4 Class I Newcastle disease viruses
2.3 遗传进化分析
根据GenBank中下载的NDV参考序列,利用MEGA软件构建系统进化树。F基因(47—420 nt)遗传进化分析结果显示,4株分离株均为Ⅰ类新城疫病毒,但不属于基因型1~10(图2)。随后,利用完整F基因编码区序列构建系统进化树[11],结果显示,4株病毒均为1c亚型,与北美分离株相似性最高(图3),虽与2015年我国广西分离株(duck/Guangxi/1261/2015)所属分支相同,但仍具有一定的差异性(图3)。
3 讨 论
3.1 Ⅰ类新城疫毒力及抗原性的演化
1990年爱尔兰暴发的新城疫是由ClassⅠ强毒株引起,分离株与该地区水鸟源无毒株亲缘关系很近,这是迄今为止唯一关于ClassⅠ新城疫病毒导致疫病流行的报道[12]。NDV最早是以弱毒形式存在,在多种压力之下才逐渐进化为强毒[13],1999—2000年,澳大利亚暴发新城疫病原株是由1998年之前当地流行的Ⅰ型弱毒株演化而来。2005年,仇旭升等[14]分离到一株基因Ⅲ型新城疫病毒,与Ⅰ系苗Mukteswar相似性在99%以上,毒力却明显增强。秦卓明等[15]以La Sota、Clone30等疫苗株和经典强毒株F48E9为对照,对国内2001—2004年间分离的13株NDV进行了鸡胚中和试验,结果表明:La Sota与Clone30和F48E9之间的中和相关指数(R)较高,但与目前流行株的中和相关指数普遍偏低。
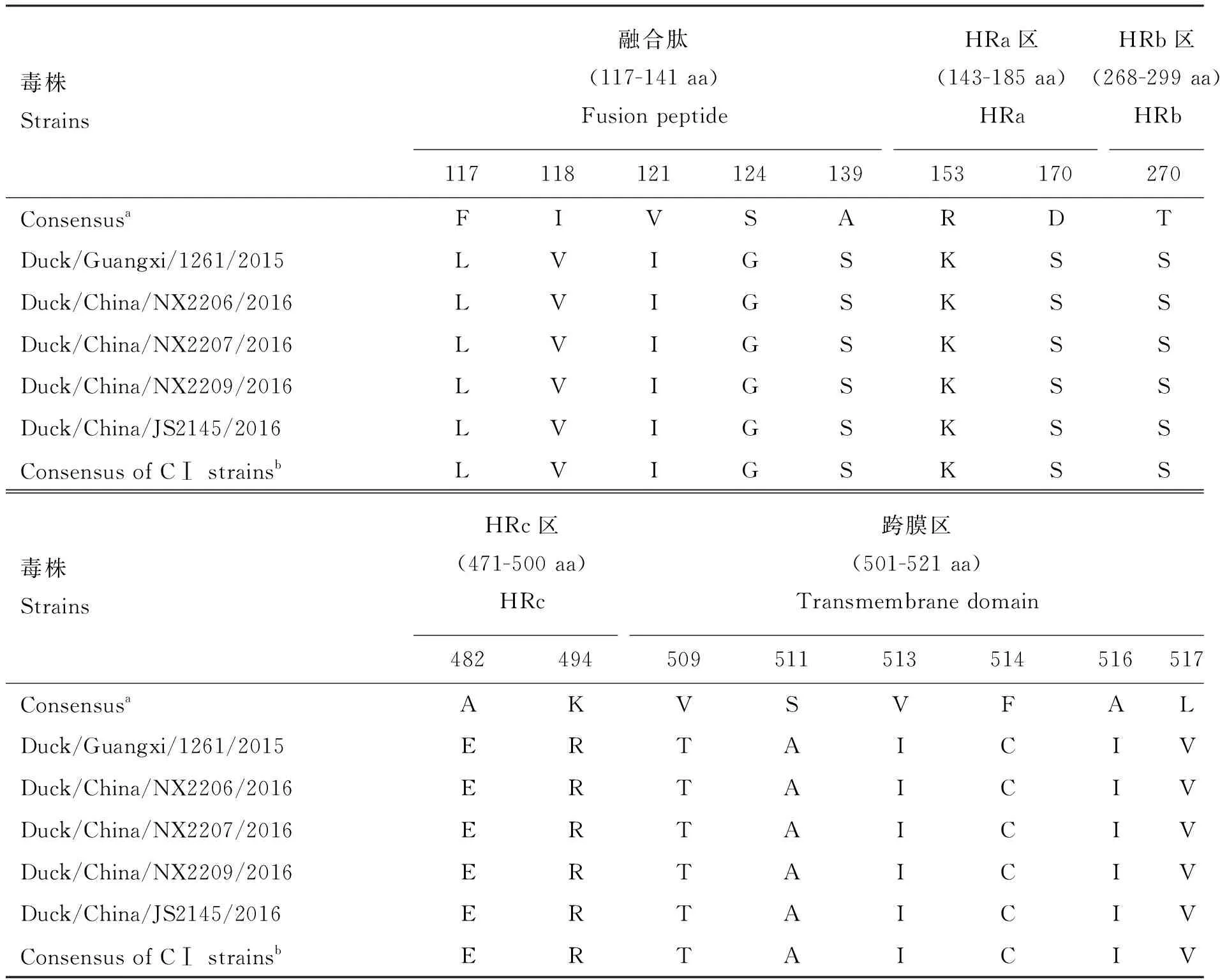
表4 I类新城疫病毒F蛋白功能区氨基酸替代分析
a. 共有序列来自不同基因型的新城疫病毒株[9];b. CⅠ毒株共有序列来自 GenBank中的40株CⅠ型的新城疫病
a. The consensus amino acid sequence was derived from NDV strains of different genotypes[9];b. Consensus of CⅠ strains was derived from 40 NDV strains of CⅠ from GenBank ClassⅠ新城疫毒株的宿主主要为易于迁徙的水禽和野鸟,容易导致NDV由野鸟或水禽向家禽中转移,进而演化为强毒株[4]。Ⅰ类新城疫病毒在我国分布广泛[3],尽管目前的监测结果证实大多数分离株存在于活禽市场等流通环节,但仍需加强活禽市场监管,强化消毒、定期休市等措施,避免新城疫病毒向养殖场传播,降低病毒返强的风险。
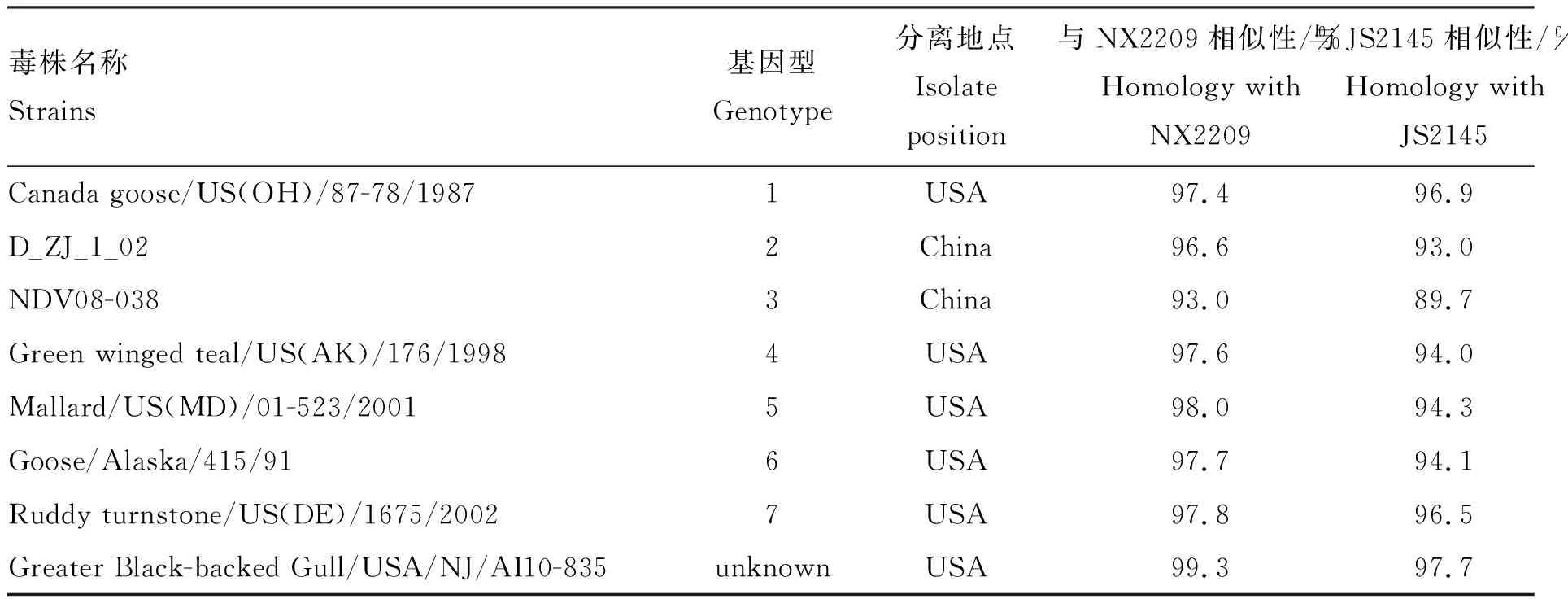
表5 新城疫病毒F蛋白氨基酸相似性比较
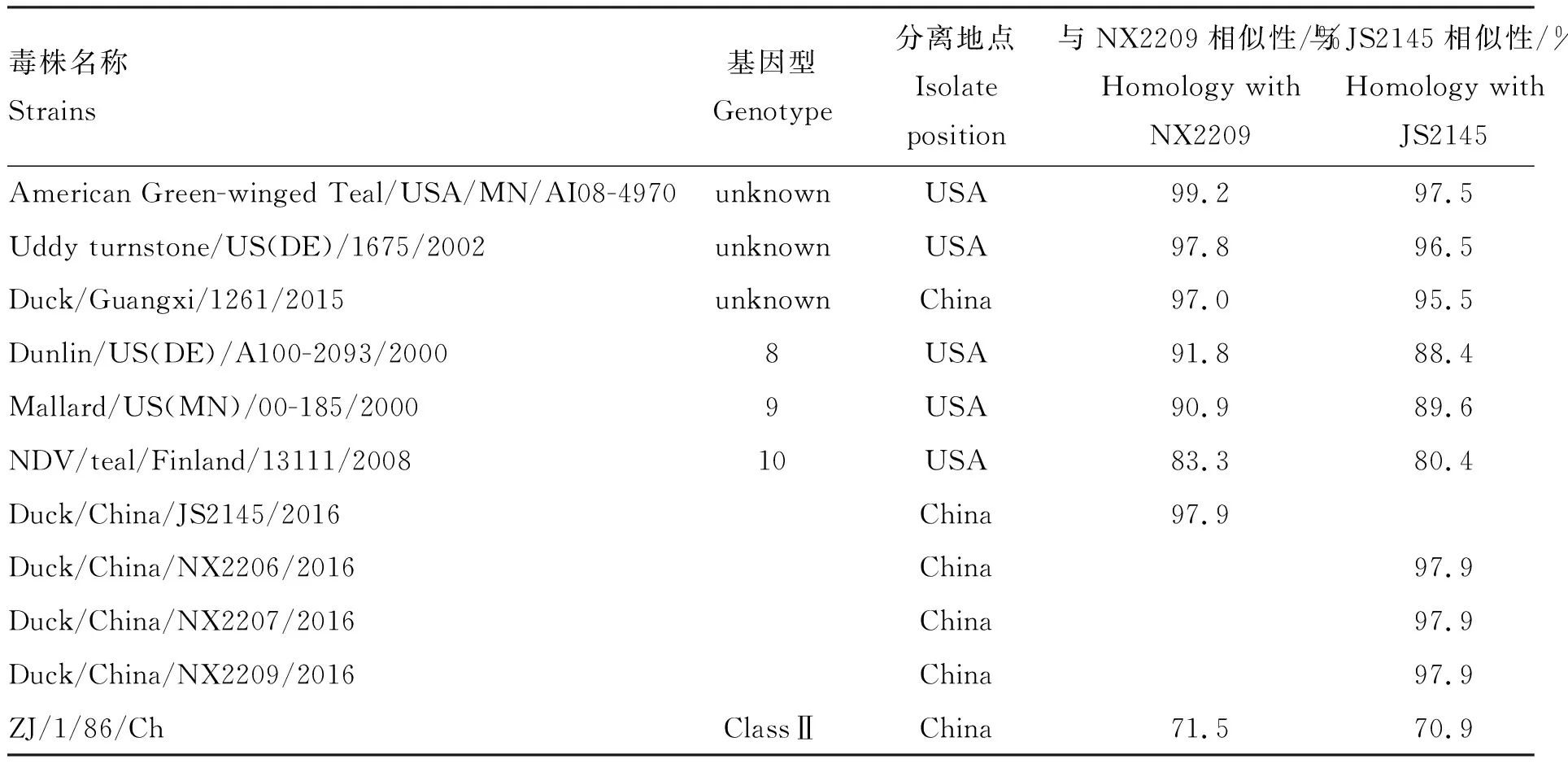
(转下页 Carried forward)
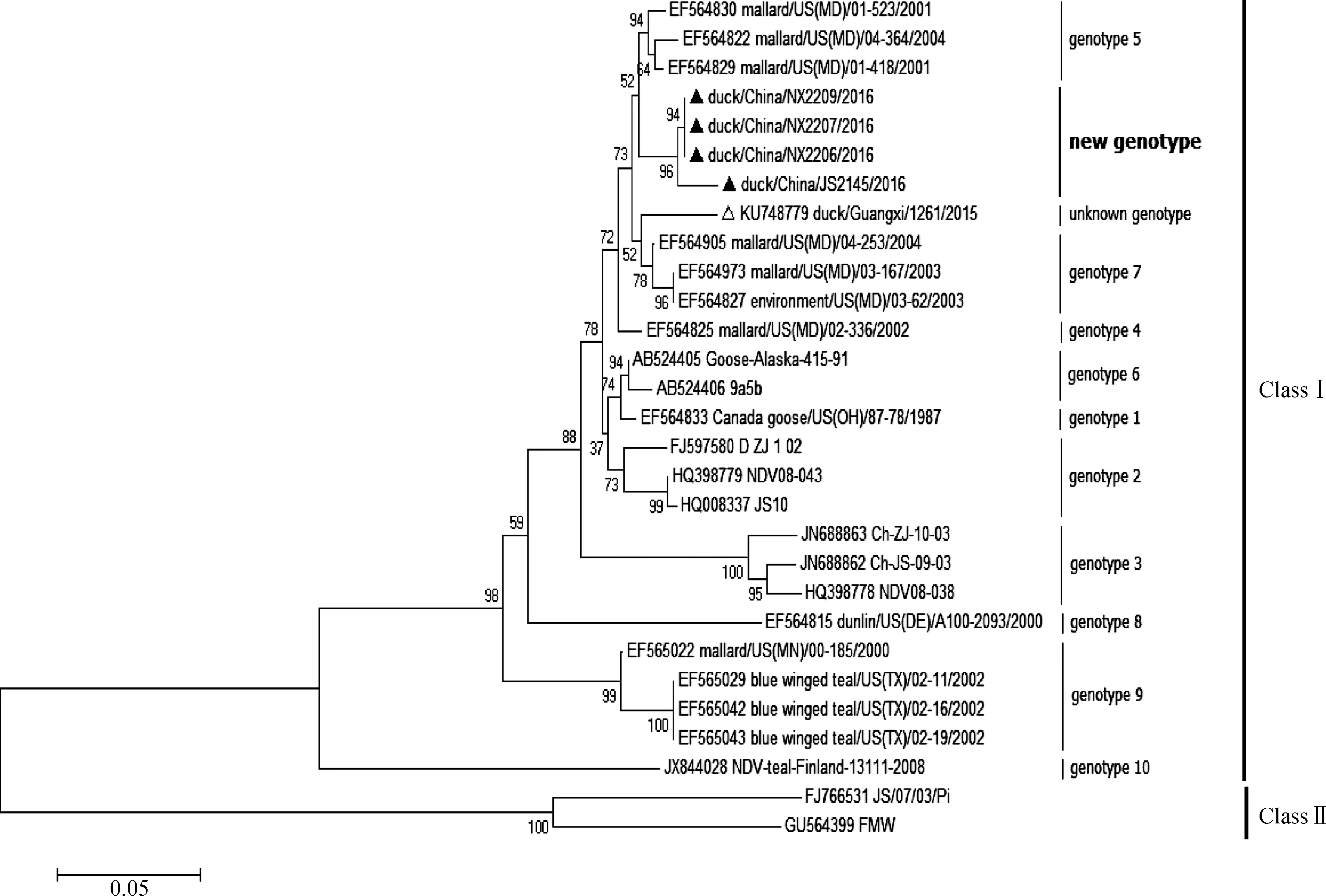
本文中的新城疫病毒分离株用▲标示,GX1261用△标示 NDV isolates used in the study were labeled by▲, GX1261 were labeled by△图2 Ⅰ类新城疫病毒F基因(47—420 nt)遗传进化树Fig.2 The phylogenetic tree of F gene(47-420 nt)among Newcastle disease virus of Class Ⅰ
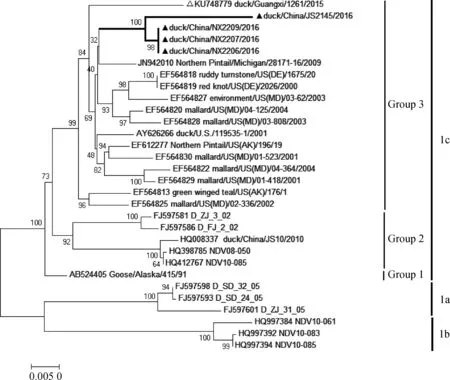
▲.新城疫分离株▲.NDV isolates used in the study图3 Ⅰ类新城疫病毒F基因遗传进化树Fig.3 The phylogenetic tree of F gene among Newcastle disease virus of Class Ⅰ
3.2 我国新出现的Ⅰ类NDV的流行现状
本研究中分离的4株Ⅰ类NDV 与北美分离株的F蛋白氨基酸相似性较高,与duck/Guangxi/1261/2015的相似性仅为95.5%~97.0%。其中分离株Duck/China/NX2206/2016、Duck/China/NX2207/2016、Duck/China/NX2209/2016 F氨基酸相似性为100%,提示这3株分离株可能有共同的来源,此3株分离株与Duck/China/JS2145/2016的氨基酸相似性为97.9%。
用F基因47—420 nt高变区序列构建系统进化树,4株病毒均属于Ⅰ类新城疫病毒,与基因5型遗传进化关系较为接近,但与基因5型代表株所在分支的bootstrap值较低,与2015年国内分离到的Ⅰ类未知基因型毒株(GX1261)在遗传关系上也存在一定差异。利用F基因编码区全长序列进行遗传进化分析,结果显示,4株病毒和2015年国内Ⅰ类分离株(GX1261)同属于基因1c亚型,与北美分离株位于同一分支(Group 3),此结果与F蛋白同源性分析结果一致。
基因分型结果说明,这4株病毒为我国新出现的Ⅰ类新城疫病毒,虽然与GX1261相似性较高,但仍存在一定差异。由于缺乏全球Ⅰ类新城疫病原学监测数据,至今无法确定此类病毒的确切来源与传播途径。根据现有数据推测,该病毒出现于我国可能与带毒候鸟的长距离迁徙有关。目前,虽然只在宁夏、江苏和广西的水禽(鸭)中分离到这种新型NDV,但水禽被认为是NDV的自然宿主,在贮存和传播病毒方面具有重要作用,而水禽和陆禽的混合饲养也给病毒的传播和扩散带来便利。因此,应加强Ⅰ类NDV的系统监测,同时避免水禽和陆禽的混合饲养,防止病原进一步扩散和毒力返强。
4 结 论
2016年分离到4株Ⅰ类新城疫病毒,其F基因编码长度均为1 662 nt,编码553个氨基酸,毒株间氨基酸相似性为97.9%~100%。分离株F蛋白裂解位点的氨基酸组成均为112ERQERL117,符合弱毒株的分子特征。F基因高变区(47—420 nt)遗传进化分析结果提示这4株病毒为国内新出现的基因型,属于基因1c亚型,与北美分离株高度同源。
