不同海拔圆叶天女花的传粉生物学研究
2018-08-30陈小红李慧敏付梦媛肖书礼
陈小红, 李慧敏, 杨 科, 付梦媛, 肖书礼
(1. 四川农业大学林学院, 四川 成都 611130; 2. 长江上游生态安全协同创新中心, 四川 成都 611130; 3. 中国科学院昆明植物研究所, 云南 昆明 650201)
植物花部特征主要包括花的结构、颜色、气味和分泌物质,还包括花的大小、开放数量和空间排列方式等[1]。作为被子植物的有性繁殖器官,花较其他繁殖器官表现出更高的变异性,在植物交配系统进化中的意义重大,影响植物的繁殖策略[2]。植物花部特征的变化不仅影响植物对传粉媒介的吸引和昆虫的传粉效率,还影响虫媒传粉植物有性繁殖的成功率及其对外界环境的适应能力[3]。随着海拔的升高,影响植物生长和繁殖的环境因子(如温度和光照)不断变化,并对植物花部特征产生一定影响。已有研究结果表明:海拔作为外界环境因子对大通翠雀花(DelphiniumpylzowiiMaxim.)[4]、狼毒(StellerachamaejasmeLinn.)[5]、露蕊乌头(AconitumgymnandrumMaxim.)[6]、风毛菊属(SaussureaDC.)[7-9]等植物的花部特征均具有显著影响。环境因子和植物花部特征的变化影响各类昆虫的活动,访花昆虫的种类和访问频率在不同生境中存在较大差异,进而影响植物的繁殖策略[10-14]。
圆叶天女花〔Oyamasinensis(Rehder et E. H. Wilson) N. H. Xia et C. Y. Wu〕隶属于木兰科(Magnoliaceae)天女花属〔Oyama(Nakai) N. H. Xia et C. Y. Wu〕,为落叶灌木或小乔木[15]。圆叶天女花仅分布于四川中部及北部海拔1 800~2 500 m的林缘或灌丛,其中,在雅安芦山和成都彭州集中分布,为国家Ⅱ级重点保护野生植物、准四川省特有植物和优先拯救保护的极小种群植物[16]。曾洪等[17]认为,圆叶天女花生态位宽度较大,生存状况良好,生态适应能力较强,在群落中具有一定种群优势。但圆叶天女花在恶劣环境的生态适应机制尚未深入研究,限制了其有效保护策略的制定。
鉴于此,作者对不同海拔居群圆叶天女花的花部特征和访花昆虫进行研究,以期阐明其花部特征和传粉生物学对海拔变化的响应,深入了解其繁殖特性和生态适应性,并为圆叶天女花的生殖生态学研究和保护利用对策提供参考。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于四川省雅安市芦山县大川镇黄水河大熊猫基因交流走廊带(地理坐标为北纬30°32′~30°49′、东经102°52′~103°11′),面积约400 hm2,地处青衣江上游,属中纬度内陆亚热带湿润气候,雨量充沛,7月至8月降水量达1 000~1 800 mm,年均气温11 ℃。土壤类型为山地黄棕壤,呈微酸性。该地植被类型为山地针阔混交林,植被分层不明显,为天然次生林。
圆叶天女花多生长在沟谷的崖壁和陡坡,生境恶劣,生境郁闭度0.5~0.7。除零星生长的铁杉〔Tsugachinensis(Franch.) Pritz.〕、卵叶钓樟(LinderalimprichtiiH. Winkl.)、领春木(EupteleapleiospermumHook. f. et Thoms.)、槭属(AcerLinn.)、柳属(SalixLinn.)、粉花绣线梅(NeilliarubifloraD. Don)、川莓(RubussetchuenensisBureau et Franch.)和贴毛箭竹(FargesiaadpressaYi)等木本植物外,圆叶天女花分布较集中,为优势种。
在对该区域圆叶天女花资源全面踏查的基础上,沿海拔梯度选取4个面积不小于10 m×40 m的自然居群,分别编号为P1、P2、P3和P4,各居群基本信息见表1。其中,圆叶天女花在P1居群中与领春木和卵叶钓樟等混生,在P2居群中与槭属和柳属等混生,在P3居群中与柳属和铁杉等混生,在P4居群中与贴毛箭竹和铁杉等混生。在各圆叶天女花居群内随机选取10株生长正常、无明显缺陷和病虫害的圆叶天女花开花植株作为花朵采样植株,各植株间距在5 m以上。
1.2 研究方法
1.2.1 花部特征测定 各圆叶天女花植株分别随机选取3朵已经盛开且雄蕊未完全散粉的花朵,每个居群共30朵,使用游标卡尺(精度0.1 mm)测量花朵的直径和长度、花被片的长度和宽度以及雄蕊和雌蕊的长度,统计雄蕊和雌蕊的数量。
表1圆叶天女花不同海拔居群的基本信息
Table1BasicsituationofpopulationsofOyamasinensis(RehderetE.H.Wilson)N.H.XiaetC.Y.Wuatdifferentaltitudes
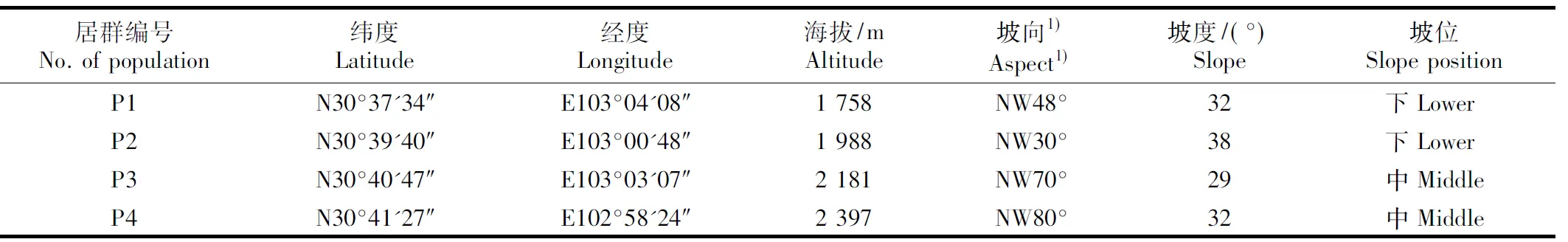
1)NW: 北偏西 North by west.
各圆叶天女花植株分别随机选取3朵即将开放的花蕾固定于FAA固定液中,然后带回实验室,每个居群共30朵。分别将每朵花的花药全部取出,用蒸馏水清洗后置于5 mL离心管中,充分摇匀后取1 μL,在BX51光学显微镜(日本Olympus公司)下观察统计花粉数,重复观察10次,计算每朵花花粉数的平均值;解剖每朵花的雌蕊群,光学显微镜下统计胚珠数。
1.2.2 柱头可授性与花粉活力检测 参考文献[14],各圆叶天女花植株分别随机选取3朵当日即将开放的花朵并标记,采集后立刻带回实验室并置于4 ℃冰箱冷藏,每个居群共30朵。采集当日及之后每隔24 h取出一批进行TTC染色,直至花粉没有活性,光学显微镜下统计3~5个视野内的染色花粉数和未染色花粉数,根据公式“花粉活力=〔染色花粉数/(染色花粉数+未染色花粉数)〕×100%”计算花粉活力。
各圆叶天女花植株分别随机选取3朵当日即将开放的花朵,将柱头浸入盛有联苯胺-过氧化氢溶液的凹面载玻片中,放大镜下观察柱头变化情况。其中,无气泡和变蓝表示柱头无可授性,有气泡产生和轻微变蓝表示柱头可授性一般,有较多的气泡和较明显的变蓝表示柱头可授性强。每天观察,直至最早标记的花的柱头完全变黑。
1.2.3 访花昆虫的观察 于2017年5月8日至10日连续3 d在圆叶天女花不同海拔居群同时观察并录像,每个居群选取2株植株,每株随机选取3朵花观察记录访花昆虫的访花行为。由于9:00之前和17:00之后基本不见访花昆虫,因此,于9:00—17:00连续观察并记录各时段访花昆虫的种类、访问次数、访问部位和访问时间,结果取平均值。同时捕捉不能识别的昆虫,并带回实验室鉴定。
1.3 数据统计和处理
采用EXCEL 2010软件对实验数据进行计算和分析,采用SPSS 20.0统计分析软件中的one-way ANOVA对圆叶天女花不同海拔居群间的差异显著性进行检验。
2 结果和分析
2.1 不同海拔居群圆叶天女花花朵和花被片的形态特征分析
圆叶天女花的花单生枝顶,花梗下弯,花朵大型,花朵直径10 cm以上,花朵长约3 cm。不同海拔居群圆叶天女花花朵和花被片的形态特征见表2。由表2可以看出:不同海拔居群间圆叶天女花的花朵直径差异明显。P2(海拔1 988 m)和P3(海拔2 181 m)居群圆叶天女花的花朵直径显著(P<0.05)大于P1(海拔1 758 m)和P4(海拔2 397 m)居群,表明海拔1 988~2 181 m区域圆叶天女花的花朵展示效果较好。不同海拔居群间圆叶天女花的花朵长差异不显著(P>0.05)。
观察结果显示:圆叶天女花的花萼与花瓣相似,同被花,花被片9,每轮3,白色。第1轮花被片为花萼,倒卵形,较短小;第2轮花被片卵圆形,大于内、外轮(即第1和第3轮)花被片;第3轮花被片椭圆形,较短小。
由表2还可以看出:总体上看,P3居群第1、第2和第3轮的花被片长和花被片宽均最大,P2居群次之,P4居群较小,P1居群最小,说明P2和P3居群圆叶天女花的花被片较大,展示效果较明显,能吸引更多昆虫到访。


居群Population花朵Flower第1轮花被片The 1st whorl of tepals第2轮花被片The 2nd whorl of tepals第3轮花被片The 3rd whorl of tepals编号No.海拔/mAltitude直径/mmDiameter长度/mmLength长度/mmLength宽度/mmWidth长度/mmLength宽度/mmWidth长度/mmLength宽度/mmWidthP11 758105.71±7.97b30.80±3.17a48.15±5.10b26.65±3.93c54.31±4.17b36.95±8.70c50.45±3.30c31.01±5.39bP21 988129.88±24.24a29.65±2.95a58.68±2.27a31.19±3.22ab67.60±7.52a50.24±6.92ab59.82±5.83a34.11±9.33bP32 181135.56±6.57a32.53±1.50a59.89±4.64a34.15±2.76a69.60±4.86a55.47±4.67a60.89±2.02a44.49±1.67aP42 397106.54±6.87b31.06±3.21a50.70±4.91b28.77±0.56bc59.88±4.33b45.82±3.91b54.47±0.17b36.27±3.47b
1)同列中不同的小写字母表示差异显著(P<0.05) Different lowercases in the same column indicate the significant difference (P<0.05).
2.2 不同海拔居群圆叶天女花雄蕊和雌蕊的特征分析
圆叶天女花为两性花,雄蕊群紫红色,离生雄蕊多数;雌蕊群浅绿色,离生心皮雌蕊多数,雄蕊群与雌蕊群间无间隔。不同海拔居群圆叶天女花雄蕊和雌蕊的特征见表3。由表3可以看出:P1(海拔1 758 m)居群的雄蕊数和花粉数均显著大于其他居群,但雄蕊长较短;P1居群的雌蕊数和胚珠数均显著大于其他居群,但4个居群间雌蕊长无显著差异。4个居群的花粉/胚珠比(P/O)为5 313~11 453,且随着海拔升高,P/O值逐渐减小,表明圆叶天女花在雄蕊和雌蕊投入上随着海拔升高而逐渐减少。圆叶天女花的P/O值符合Cruden[18]的标准,表明其为专性异交,需要传粉者。

居群 Population雄蕊 Stamen编号No.海拔/mAltitude数量Number长度/mmLength花粉数Pollen number雌蕊 Pistil数量Number长度/mmLength胚珠数Ovule number花粉/胚珠比Pollen/ovule ratioP11 75883.5±10.5a10.5±1.2c938 000±325a41.5±4.3a4.0±1.1a81.9±1.4a11 453P21 98878.3±10.7b13.2±1.7b629 300±252b32.6±2.3b3.9±0.6a64.5±1.0b9 757P32 18177.8±4.6b14.8±0.7a537 000±187c30.4±2.9b4.1±0.1a59.5±0.8c9 025P42 39766.0±11.4c10.1±0.6c349 600±248d33.3±2.1b4.7±0.6a65.8±0.8b5 313
1)同列中不同的小写字母表示差异显著(P<0.05) Different lowercases in the same column indicate the significant difference (P<0.05).
2.3 不同海拔居群圆叶天女花的花粉活力和柱头可授性分析
不同海拔居群圆叶天女花的花粉活力和柱头可授性的动态变化见表4。由表4可以看出:不同海拔居群圆叶天女花的花粉活力变化趋势基本一致,均随开花时间的延长逐渐降低。开花第2至第4天,花粉活力较高,总体在60%以上,之后花粉活力明显下降,到开花第6天花药脱落,再无花粉散出。总体上看,同一开花时间P4(海拔2 397 m)居群的花粉活力最高,P3(海拔2 181 m)居群的花粉活力次之,P2(海拔1 988 m)居群的花粉活力较低,P1(海拔1 758 m)居群的花粉活力最低,且P4和P3居群在不同开花时间的花粉活力总体上显著高于P2和P1居群。其中,P4居群开花第1天的花粉活力最高(93.1%),开花第5天时花粉活力仍保持在50%以上,表明该居群的花粉活力保存时间较长。P1居群的花粉活力最低(开花第2天时花粉活力为85.7%),且随开花时间延长花粉活力下降最快,开花第5天花粉活力仅20.9%。
开花第1天,4个居群圆叶天女花的柱头上均有粘液分泌,放大镜下可见柱头周围呈蓝色,且有大量气泡产生,表明此时柱头可授性强,但此时并无花粉散出。开花第2天,4个居群的花药开裂花粉散出,部分柱头开始出现褐变斑点,可授性变弱。开花第3天,P1和P2居群的柱头内向贴合,无粘液分泌,无可授性;而P3和P4居群的柱头仍具有可授性,但可授性一般。开花第4天及之后,4个居群的柱头均无可授性。



居群 Population不同开花时间的花粉活力/%1) Pollen viability at different flowering times1)编号No.海拔/mAltitude第1天The 1st day第2天The 2nd day第3天The 3rd day第4天The 4th day第5天The 5th day第6天The 6th dayP11 758*85.7±1.9b80.5±8.9c63.4±4.8b20.9±3.9c*P21 988*86.0±2.6b83.0±4.9b58.6±4.6c40.8±8.2b*P32 181*91.6±1.1a84.2±0.9a65.5±2.5b55.8±2.4a*P42 397*93.1±1.0a85.6±2.0a75.8±1.4a57.2±5.3a*居群 Population不同开花时间的柱头可授性2) Stigma receptivity at different flowering times2)编号No.海拔/mAltitude第1天The 1st day第2天The 2nd day第3天The 3rd day第4天The 4th day第5天The 5th day第6天The 6th dayP11 758+++/-----P21 988+++/-----P32 181+++/-+/----P42 397+++/-+/----
1)*: 未检测到花粉Pollen undetected. 同列中不同的小写字母表示差异显著(P<0.05) Different lowercases in the same column indicate the significant difference (P<0.05).
2)++: 柱头可授性强Strong stigma receptivity; +/-: 柱头可授性一般Common stigma receptivity; -: 柱头无可授性Stigma without receptivity.
2.4 不同海拔居群圆叶天女花访花昆虫的种类及访花行为分析
5月8日至5月10日,不同海拔居群圆叶天女花访花昆虫的种类及访花行为见表5。由表5可以看出:4个居群圆叶天女花的访花昆虫共13种,访问次数共317次,其中膜翅目(Hymenoptera)和双翅目(Diptera)昆虫的访问次数较多。其中,熊蜂(Bombusningpoensis)访问次数为171次,占访花昆虫总访问次数的53.9%,其访问部位主要为花被片、雄蕊和雌蕊,每朵花访问时间为(44.7±4.8) s。花蚊子(Aedesalbopictus)和中华蜜蜂(Apiscerana)的访问次数分别为52和38次,在花朵的各个部分均有停留,在雄蕊和雌蕊间来回穿梭,每朵花访问时间几秒到几十秒不等,是圆叶天女花的有效传粉昆虫。食蚜蝇(Syrphussp.)访问次数较少,为15次,但每朵花访问时间长,为(367.0±107.8) s,主要停留在花被片、雄蕊和雌蕊上,也是有效传粉昆虫。白粉蝶(Pierisrapae)访问次数也较少,仅5次,访问部位为雌蕊和雄蕊,能有效传递花粉。蚁科(Formicidae)、寄蝇科(Tachinidae)、叶甲科(Chrysomelidae)、花金龟科(Cetoniidae)和丽蝇科(Calliphoridae)访花昆虫的访问次数较少,且仅停留在花被片上,不计为有效传粉昆虫。圆叶天女花花朵中花蓟马(Frankliniellaintonsa)数量较多,主要取食花被片、雄蕊和雌蕊,破坏了花朵的完整性,导致花朵无法正常结实,其传粉性无法估计。
表5不同海拔居群圆叶天女花访花昆虫的种类及访花行为(5月8日至5月10日)
Table5Speciesandvisitingbehaviorsofflower-visitinginsectsofOyamasinensis(RehderetE.H.Wilson)N.H.XiaetC.Y.Wuinpopulationsatdifferentaltitudes(fromMay8toMay10)

目Order科Family种类Species访问部位1)Visiting part1)不同居群的访问次数2)Visiting times in different populations2)P1P2P3P4膜翅目Hymenoptera熊蜂科Bombidae熊蜂Bombus ningpoensisTe,St,Pi71433819蜜蜂科Apidae中华蜜蜂Apis ceranaTe,St,Pi4121012蚁科Formicidae小黄蚁Monomorium pharaonisTe2112双翅目Diptera蚊科Culicidae花蚊子Aedes albopictusTe,St,Pi2412106寄蝇科Tachinidae园寄蝇Paradejeania sp.Te0213食蚜蝇科Syrphidae食蚜蝇Syrphus sp.Te,St,Pi6540头蝇科Pipunculidae头蝇Pipunculus sp.Te8000鞘翅目Coleoptera叶甲科Chrysomelidae杨叶甲Chrysomela populiTe0120花金龟科Cetoniidae花金龟Cetonia sichuanaTe0001鳞翅目Lepidoptera丽蝇科Calliphoridae红头丽蝇Calliphora vicinaTe0100凤蝶科Papilionidae白粉蝶Pieris rapaeSt,Pi0320蜘蛛目Araneae蟹蛛科Thomisidae三突花蛛Misumenops tricuspi-datusTe0470缨翅目Thysanoptera蓟马科Thripidae花蓟马Frankliniella intonsaTe,St,Pi****
1)Te: 花被片Tepal; St: 雄蕊Stamen; Pi: 雌蕊Pistil.
2)P1,P2,P3,P4: 分别为海拔1 758、1 988、2 181和2 397 m处的圆叶天女花居群RepresentingOyamasinensis(Rehder et E. H. Wilson) N. H. Xia et C. Y. Wu populations at altitudes of 1 758, 1 988, 2 181 and 2 397 m, respectively. *: 花解剖时发现大量花蓟马,但未观测到其访花行为,访问次数无法统计ManyFrankliniellaintonsaare found during flower anatomy, their visiting behaviors are not observed, so their visiting times are uncounted.
由表5还可以看出:不同海拔居群圆叶天女花访花昆虫的种类也存在较大差异。P1(海拔1 758 m)和P4(海拔2 397 m)居群的访花昆虫种类均为7种,由于P2(海拔1 988 m)和P3(海拔2 181 m)居群圆叶天女花花朵的展示效果明显,能吸引更多的访花昆虫到访,如白粉蝶和三突花蛛(Misumenopstricuspidatus)等,访花昆虫种类为10~11种。从访问次数来看,随着海拔的升高,P1至P4居群访花昆虫的访问次数逐渐减少。其中,P1居群访花昆虫的总访问次数显著高于其他居群,总访问次数为115次,平均每小时访问14.4次;P4居群访花昆虫的总访问次数仅43次,平均每小时访问5.4次。
5月9日不同海拔居群不同时段熊蜂访问圆叶天女花花朵的次数见图1。由图1可以看出:9:00—17:00,P1居群熊蜂访问圆叶天女花花朵的次数呈“减少—增加—减少”的变化趋势,其中,在11:00—15:00间各时段的访问次数较多(大于等于9次);P2居群熊蜂访问圆叶天女花花朵的次数呈“增加—减少—增加—减少”的变化趋势,分别在10:00—11:00和13:00—14:00时段达到峰值,访问次数分别为9和10次;P3和P4居群熊蜂访问圆叶天女花花朵的次数总体上呈先增加后减少的变化趋势,均在10:00—11:00时段达到峰值,访问次数分别为10和9次。在10:00—14:00间的各时段,4个居群中熊蜂的总访问次数较高。其中,在11:00—15:00间各时段,P1居群熊蜂的访问次数较多,均在9次以上,而P3和P4居群熊蜂的访问次数均在5次以下。

—○—: P1居群(海拔1 758 m)Population P1 (altitude of 1 758 m); —●—: P2居群(海拔1 988 m)Population P2 (altitude of 1 988 m); —△—: P3居群(海拔2 181 m)Population P3 (altitude of 2 181 m); —▲—: P4居群(海拔2 397 m)Population P4 (altitude of 2 397 m).Ⅰ: 9:00—10:00; Ⅱ: 10:00—11:00; Ⅲ: 11:00—12:00; Ⅳ: 12:00—13:00; Ⅴ: 13:00—14:00; Ⅵ: 14:00—15:00; Ⅶ: 15:00—16:00; Ⅷ: 16:00—17:00.图1 不同海拔居群不同时段熊蜂访问圆叶天女花花朵的次数(5月9日)Fig. 1 Visiting times of Bombus ningpoensis on flowers of Oyama sinensis (Rehder et E. H. Wilson) N. H. Xia et C. Y. Wu in different time segments in populations at different altitudes (May 9)
3 讨 论
植物的花部特征与环境因子之间有着密切关联,因生境差异其构件在形态上存在差异,从而影响植物的繁殖特征[2]。对大通翠雀花[4]和风毛菊属[7-9]等植物的研究结果表明:花被片大小以及花蕊的大小和数量均与海拔呈正相关,花粉数与海拔呈负相关,总体上看,随着海拔升高,植物的繁殖投入逐渐增加。随着海拔升高,全缘叶绿绒蒿〔Meconopsisintegrifolia(Maxim.) French.〕的花部展示投入减少,但雌雄功能的投入相对稳定[14]。本研究中,随着海拔的升高,圆叶天女花花朵的各个构件在资源分配上发生了变化。P2(海拔1 988 m)和P3(海拔2 181 m)居群的花被片具有最佳展示效果,P1(海拔1 758 m)居群的雄蕊和雌蕊在大小和数量方面占有优势,P4(海拔2 397 m)居群的花粉活力和柱头可授性最好。说明随着海拔升高,圆叶天女花雄蕊和雌蕊的功能投入逐渐减少,但雄蕊和雌蕊的寿命延长,这与草本植物[4,9,13-14]有所差异,其原因可能在于圆叶天女花特殊的花部结构,圆叶天女花花梗下弯,花朵朝地面方向开放,雄蕊群处于花托上方,这一特征为雄蕊和雌蕊延长寿命提供了可能。
圆叶天女花为虫媒花,不同海拔居群访花昆虫的种类有所差异,P2和P3居群的访花昆虫种类多于P1和P4居群,P2和P3居群的花朵展示效果也最好,花被片较大以及花被片颜色为白色,在高山生境中非常醒目,有利于吸引更多的昆虫传粉,这与大多数虫媒植物[19-22]的传粉生物学特征一致。圆叶天女花的访花昆虫中,熊蜂的访问次数最多,这与高山环境下熊蜂具有较高的热调节能力、花粉传递效率以及逆境生存能力有关[23]。
昆虫的访花行为与温度密切相关,温度限制导致其在不同海拔地区的访问次数存在差异[24-25]。本研究中,随着海拔升高,气温逐渐降低,各居群访花昆虫的总访问次数和主要传粉昆虫熊蜂的访问次数均逐渐减少。由于研究地位于华西雨屏区,几乎每天14:00后都有小雨,且气温开始下降,访花昆虫活动也相应减少。而主要传粉者熊蜂的访问高峰期为10:00—14:00,该时段阳光照射最强且气温最高,有利于昆虫传粉。在降雨过程中,圆叶天女花花被片呈半合拢杯状,这种特殊现象有利于保护花蕊,但访花昆虫较少,这与单性木兰〔Woonyoungiaseptentrionalis(Dandy) Y. W. Law〕的繁殖策略相似[26]。
钟云芳等[13]对不同海拔海南凤仙花(ImpatienshainanensisY. L. Chen)花粉活力的研究结果显示:高海拔种群在开花第1至第4天的花粉活力显著低于低、中海拔种群。本研究中,圆叶天女花的花粉活力随海拔升高而增强,高海拔P4居群圆叶天女花的花粉活力在散粉期间最高,弥补了其在雄蕊的长度和数量以及花粉数上的不足,增强了自身的传粉特性。高海拔居群(P3和P4居群)的柱头可授性较低海拔居群(P1和P2居群)多持续1 d,这为高海拔居群圆叶天女花在雌蕊数量少、传粉昆虫少的情况下提供了更长的传粉时间。随着海拔升高,圆叶天女花在雄蕊和雌蕊性特征方面的资源投入越来越少,访花昆虫的访问次数也逐渐减少,但较长时间的花粉活力与柱头可授性可以减小访花昆虫活动少的不利影响,从而增强自身适应性。说明延长花粉和柱头寿命是高海拔濒危植物增强生态适应性的一种有效策略[27-29]。
综上所述,圆叶天女花在不同海拔的繁殖策略不同,展现出较强的适应能力。但本研究仅从其花部特征和访花昆虫等方面分析了其适应性,未深入分析其濒危机制,今后应加强其结实特性、种子萌发特性、生理特征和种群更新等方面的研究,为保护这一极小种群植物提供更多的科学依据;此外,还应加强其野生资源保护与种苗人工培育,提出合理的保育措施,进一步提高其种群面积和数量。
