长白山不同海拔白桦幼苗移栽至同一生境的光合及反射光谱特性
2018-08-29周驿之程艳霞
周驿之, 程艳霞,*, 樊 莹, 易 磊
1 北京林业大学理学院,北京 100083 2 北京林业大学林学院,北京 100083
海拔是影响植物生长发育、物质代谢、结构和功能等重要的生态因素之一[1- 2],海拔梯度升高带来的平均气温下降、大气压及CO2分压下降、辐射增强等对植物叶片的生理性状有重要影响[3]。不同海拔梯度上植物生理性状的变化国内外已有学者进行大量研究,如叶片表皮功能性状在海拔梯度下受环境的影响[4]、植物随海拔变化的生理适应[5]、树木的林冠结构由于海拔梯度上光照的变化导致差异性[6]等。但此类研究是基于自然状态下研究物种对环境的适应性。也有学者通过移植的方法研究植物生理性状的变化,如通过海拔互换的方式研究台湾赤杨的光合作用和水分利用率[7],通过同质园(Common garden)实验研究北美黄杉木幼苗生理和形态的改变[8],但这些研究都是在人工控制环境的条件下进行,在自然条件下进行移栽以研究物种生理性状的改变尚有不足之处。如今通过移植的技术手段研究植物生理性状的方法日趋成熟[9-10],长白山不同海拔群落生理性状变化明显,在长白山海拔梯度条件下进行幼苗移植以研究其生理性状的变化具有重要的生态学意义。
白桦(Betulaplatyphylla)作为先锋种植物,具有良好的环境适应能力[11]。白桦在长白山分布广泛,是原生红松针阔混交林受到火灾等干扰后形成的次生植被,垂直分布范围在海拔700—1700 m之间[12]。目前关于白桦的群落结构和生理特性已有研究[9,12],不同海拔来源的白桦其生理性状的差异性具体表现在叶片的各项生理指标上[13]。本研究将分布在高、中、低3个海拔梯度的白桦幼苗移栽到同一个海拔梯度上,并与原生境野生白桦幼苗进行对比,以研究白桦幼苗进入新环境后生理性状的变化,探讨白桦幼苗生理性状对环境的响应机制,以期为长白山地区白桦幼苗的发育以及森林的演替提供理论依据。
1 材料与方法
1.1 研究区域概况
本研究在长白山北坡和中国科学院长白山森林生态系统定位站内进行。长白山气候复杂,属于典型的温带大陆性山地气候,夏季高温多雨,冬季低温少雨,年平均温度5.5℃,月平均温度最低-13.7℃,最高20.4℃,年平均降水量在600—900 mm之间(图1)。长白山植被随着海拔升高,依次为夏绿阔叶蒙古栎林带(在450 m以下)、红松针阔混交林(海拔450—1000 m)、暗针叶林带(北坡海拔1000—1800 m)、岳桦林带(北坡海拔1800—1900 m)、高山苔原带(海拔1900—2300 m)和高山荒漠带(海拔2300 m以上)。

图1 长白山2016年5月—2017年5月平均气温与相对湿度Fig.1 The average temperature and relative humidity from May 2016 to May 2017 of Changbai mountain
1.2 研究方法
1.2.1 白桦幼苗移植
本研究于2016年5月进行移苗,长白山野生白桦幼苗分布广泛,易于选取足够合适的移植材料。本研究分别从长白山东北亚植物园(海拔750 m)、峡谷浮石林(海拔1200 m),双目峰线(海拔1400 m)每个海拔选取9株3—5年生株高(32.5±6.2) cm,基径(3.1±0.4) mm白桦幼苗(根据海拔高中低以下分别简称白桦幼苗BH、BM、BL),幼苗生长健康,无明显损害(表1)。幼苗采挖自3个海拔的道边,采挖后放到直径20 cm的营养钵中,带土坨移栽,将待移植的幼苗放置到海拔750 m的中国科学院长白山森林生态系统定位站(海拔750 m,42°24′2″N,128°6′29″E)站内林下进行缓苗处理,早晚各浇一次水,保证幼苗不失水萎蔫。
缓苗一星期后进行移栽,移栽地点选在中科院定位站林下,开辟3块1.5 m × 1.5 m的小样方,每个样方移栽不同海拔来源的白桦幼苗,幼苗之间彼此间隔半米,每周一次除去样方内杂草,控制林下郁闭度尽量相同。同时在站内寻找10株生长环境相似的野生白桦作为空白对照(CK)。实验地地上植被主要为蒙古栎(QuercusmongolicaFisch. ex Ledeb)、山杨(Pobulusdavidiana)、青楷槭(AcertegmentosumMaxim.)、色木槭(AcermonoMaxim.)和紫椴(TiliaamurensisRupr.),郁闭度0.59。移栽初期(T1)测量各项生理指标,经过一个生长季后(T2)于2017年6月再次进行各项生理指标测定。

表1 取样点海拔、地理坐标及群落类型
1.2.2 不同海拔来源白桦幼苗光谱数据测量
本研究使用Unispec-SC光谱仪(美国PPS公司)测量叶片光谱反射率,测量时卤光灯光强设定为100%,整合时间为4 ms,重复扫描次数为3次,叶夹上的光纤探头与叶片表面呈60°夹角,每10分钟参比一次。选取每个海拔来源白桦移栽幼苗和野生幼苗3株,每株选取10片成熟、健康的叶片进行测量,每片叶子测3次,于2016年6月8日、2017年6月10日晴天中午进行。
1.2.3 不同海拔来源白桦幼苗光响应曲线测量
在进行光谱数据测量的同时,选择各个海拔的移栽植株3—5株,每棵植株分别选取3—5片完整叶片,用便携式气体交换系统(Li- 6400, Li-Cor Inc.,Lincoln, NE, USA)对各幼苗叶片的光响应曲线进行测定,分4个晴朗无风日(2016年6月8日、9日、10日、11日和2017年6月10日、11日、13日、14日)进行测定,测量时间为上午9:30—11:30,为保持观测过程中其他环境因子稳定且适宜,设置流速为500 mL/min,气体交换室温度为25℃,相对湿度60%,使用液态标准CO2气瓶设制CO2浓度为400 μmol/mol,测量前对每个树种进行预实验,得到其大致的饱和光强,以确定诱导光强和设定光强(PPFD)梯度。使用Li- 6400的人工光源(LI- 6400-02B红蓝光源),并手动设置光强(1500、1200、1000、800、600、400、200、150、100、50、20、10、5、0 μmol m-2s-1)。
1.2.4 温度、湿度、光照和土壤养分的研究方法
2016年6月10—20日,利用HOBOwarePro全天候气候自动记录仪测定海拔750 m、1200 m和1400 m的气象数据,包括温度、湿度等,最终数据用以上10天的平均测量值。用便携式气体交换系统(Li- 6400, Li-Cor Inc.,Lincoln, NE, USA)测定3个海拔7:00—17:00的瞬时光合有效辐射。在3个海拔的待移栽幼苗4个方位上用土钻取土,土壤深度为20 cm。土壤处理:首先,将每个海拔的土壤集中到一起充分混匀;其次,混匀后的土壤样品根据四分法带回实验室,将土壤样品充分烘干;最后,采用常规方法测量各海拔土壤的元素含量。
1.3 数据处理
光谱仪测得的数据利用Multispec 5.1软件处理,计算出光谱反射率Rλ和光谱指数,其中Rλ是叶片反射与参比反射的比值。常用的植被指数之一为植被归一化指数(Normalized difference vegetation index,NDVI),这个指数与叶绿体含量有很好的线性关系,公式为
NDVI=(R750-R675)/(R750+R675)[14];叶绿素归一化指数(Chlnormalized difference index,ChlNDI)是对叶绿素的反应,通过ChlNDI=(R750-R705)/(R750+R705)[15]求得;类胡萝卜素指数(Carotenoid reflectance index,CRI)反映波长550 nm 和700 nm处反射值倒数与510 nm处反射值倒数的差异,该指数对类胡萝卜素的变化更为敏感,公式为CRI=R800×(1/R550-1/R700)[16];与结构无关色素指数(Structure-independent pigment index,SIPI)反映出单位叶绿素的类胡萝卜素含量,公式为SIPI=(R800-R445)/(R800+R445)[17];光化学反射指数(Photochemical reflectance index,PRI)与单位叶绿素的类胡萝卜素含量有很高的相关性,公式为PRI=(R531-R570)/(R531+R570)[18]。同时求出光谱反射曲线的一阶微分, 680—750 nm间曲线最大值的波长位置即红边位置,用红边位置(λRE,red edge)评价叶绿素含量,公式为λRE=(Ri+1-Ri-1)/Δλ[19]。
光响应曲线用Photosyn 3软件进行拟合,得到拟合曲线以及表观量子效率(Apparent quantum yield,AQE)、最大净光合速率(Maximum net photosynthetic rate,Amax)、光补偿点(Light compensation point,LCP)、光饱和点(Light saturation point, LSP)、暗呼吸速率(Dark respiration rate,Rd)和判定系数R2;同时得到光饱和条件下白桦幼苗叶片蒸腾速率(Transpiration rate,Tr, mmol m-2s-1)、气孔导度(Stomatal conductance,Gs, mol m-2s-1) 、胞间CO2浓度(Intercellular CO2concentration,Ci, μmol/mol)、叶面饱和蒸气压亏缺(Leaf saturated vapor pressure deficit, VPD, Pa)等生理因子。
白桦光谱数据以及光响应曲线数据均用R语言进行分析并绘制图表,采用单因素方差分析方法(one-way ANOVA)比较不同海拔梯度来源白桦幼苗叶片光合生理性状之间的差异,采用t-检验分析有差异的变量间的差异显著性。
2 结果与分析
2.1 不同海拔温度、湿度、光合有效辐射及土壤元素含量的变化
2016年6月份,海拔750 m处的最高温度为24.96℃,昼夜温差为13.2℃;海拔1200 m处的最高温度为22.79℃,昼夜温差为13.03℃;海拔1400 m处的最高温度为21.13℃,昼夜温差为13.04℃,最高温度均出现在12:00时左右。而湿度的变化与温度正好相反,最高湿度出现在凌晨3时左右,海拔750 m处的最高湿度为75.19%,海拔1200 m处的最高湿度为84.21%,海拔1400 m处的最高湿度为76.68%,相较而言海拔750 m 处的湿度变化较大。随着海拔升高,瞬时光合有效辐射逐渐增强(图2)。

图2 长白山2016年6月不同海拔温度、湿度和光照的比较Fig.2 Temperature, humidity and radiation at different altitude in June of Changbai mountain
不同海拔土壤之间的pH值和土壤元素含量差异较大,海拔1200 m处土壤的pH值最低,土壤有效氮含量也相应较低;海拔750 m处取样点与移栽的实验点之间土壤pH与有效氮含量差异不显著(P>0.05)(表2)。

表2 不同海拔取样点及实验地土壤pH值及营养含量
2.2 不同海拔来源白桦幼苗光响应曲线比较
2.2.1 不同海拔来源白桦幼苗移栽前后光响应曲线的变化
图3为移栽初期(T1)和经过一个生长季后(T2)6种处理白桦幼苗的光响应曲线。6种白桦幼苗的净光合速率(Pn)在PAR小于50 μmol m-2s-1条件下,白桦幼苗的光响应曲线变化规律均无显著性差异,而在PAR大于50 μmol m-2s-1条件下出现显著差异(P<0.05)。由表3可知,移栽初期(T1)不同海拔原生幼苗各项生理性状存在差异,不同海拔来源的白桦移栽幼苗各项生理性状之间也存在差异。移栽初期野生白桦最大净光合速率(Amax)大小顺序为: CK3 > CK2 > CK1,移栽幼苗最大净光合速率大小顺序为: BM>BH>BL。除了BM与BH差异不显著外(P>0.05),来源于海拔750 m的BL和CK1与BM、BH之间差异均显著(P<0.05)。经过一个生长季后,不同海拔来源的3种移栽幼苗(BH、BM、BL)与海拔750 m的野生幼苗(CK1)之间最大净光合速率(Amax)虽然存在差异,但差异不显著(P>0.05),并且生长季初期4种白桦幼苗的光补偿点(LCP)和暗呼吸速率(Rday)存在显著差异(P<0.05),而经过一个生长季后差异性变得不显著(P>0.05)。

图3 不同海拔来源白桦幼苗2016年移栽初期(T1)和2017年一个生长季后(T2)光响应曲线Fig.3 Light response curve of Betula platyphylla seedlings under different gradient of light intensity in 2016 (T1) and 2017 (T2)CK1:海拔750 m野生白桦幼苗 Feral Betula platyphylla seedlings from altitude of 750 m;CK2:海拔1200 m野生白桦幼苗 Feral Betula platyphylla seedlings from altitude of 1200 m;CK3:海拔1400 m野生白桦幼苗 Feral Betula platyphylla seedlings from altitude of 1400 m;BL:海拔750 m移栽幼苗 Transplanting Betula platyphylla seedlings from altitude of 750 m;BM:海拔 1200 m移栽幼苗Transplanting Betula platyphylla seedlings from altitude of 1200 m;BH:海拔1400 m 移栽幼苗 Transplanting Betula platyphylla seedlings from altitude of 1400 m
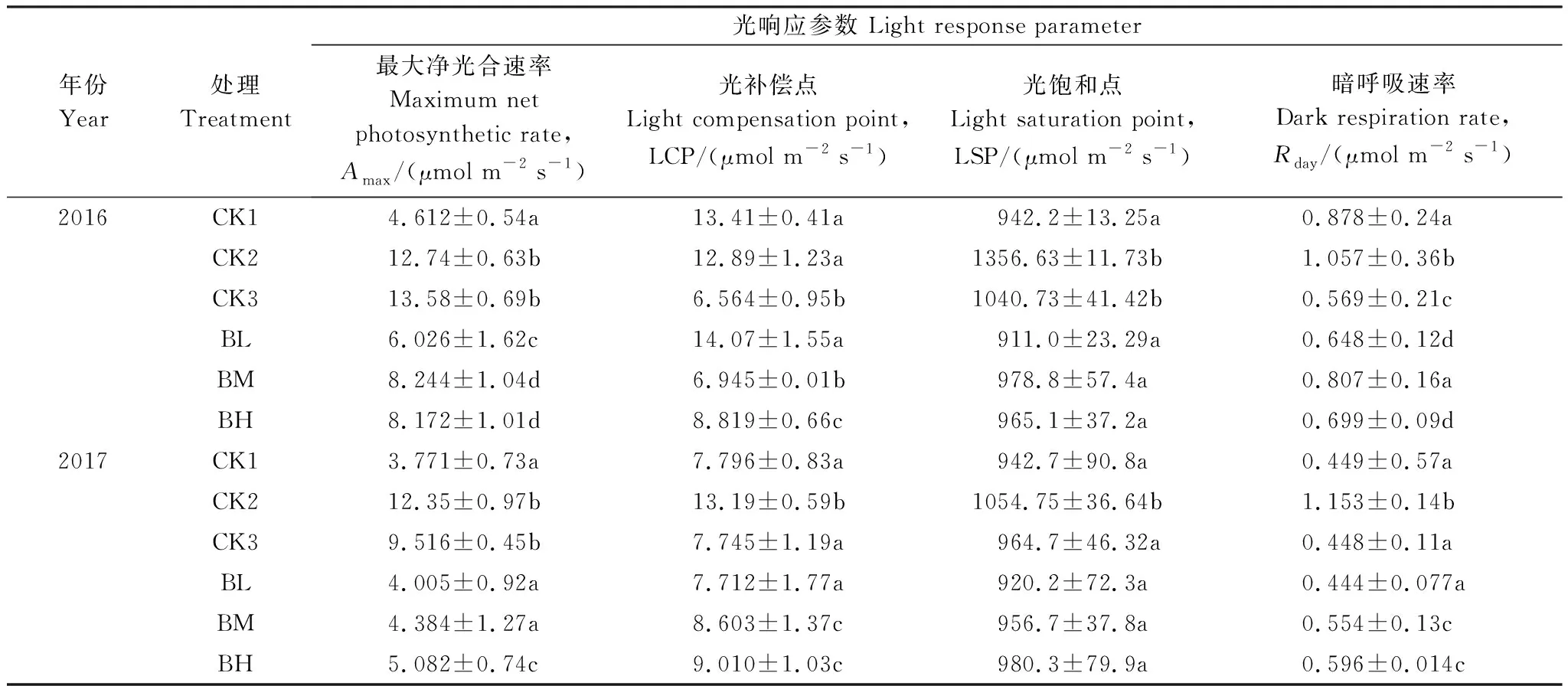
年份Year处理Treatment光响应参数 Light response parameter最大净光合速率Maximum net photosynthetic rate,Amax/(μmol m-2 s-1)光补偿点Light compensation point,LCP/(μmol m-2 s-1)光饱和点Light saturation point,LSP/(μmol m-2 s-1)暗呼吸速率Dark respiration rate,Rday/(μmol m-2 s-1)2016CK14.612±0.54a13.41±0.41a942.2±13.25a0.878±0.24aCK212.74±0.63b12.89±1.23a1356.63±11.73b1.057±0.36bCK313.58±0.69b6.564±0.95b1040.73±41.42b0.569±0.21cBL6.026±1.62c14.07±1.55a911.0±23.29a0.648±0.12dBM8.244±1.04d6.945±0.01b978.8±57.4a0.807±0.16aBH8.172±1.01d8.819±0.66c965.1±37.2a0.699±0.09d2017CK13.771±0.73a7.796±0.83a942.7±90.8a0.449±0.57aCK212.35±0.97b13.19±0.59b1054.75±36.64b1.153±0.14bCK39.516±0.45b7.745±1.19a964.7±46.32a0.448±0.11aBL4.005±0.92a7.712±1.77a920.2±72.3a0.444±0.077aBM4.384±1.27a8.603±1.37c956.7±37.8a0.554±0.13cBH5.082±0.74c9.010±1.03c980.3±79.9a0.596±0.014c
2.2.2 不同海拔来源白桦幼苗光饱和条件下生理因子比较
由表4可得,移栽初期(T1)不同海拔来源白桦幼苗在光饱和条件下各项生理因子差异性显著(P<0.05),如来源于海拔750 m的移栽(BL)和野生(CK1)白桦幼苗的蒸腾速率(Tr)和气孔导度(Gs)与来源于海拔1200 m(BM)和1400 m(BH)的白桦幼苗差异显著(P<0.05),而来源于海拔1200 m白桦幼苗(BM)的胞间CO2浓度(Ci)、水分利用率(WUE)和叶面饱和蒸气压亏缺(VPD)等生理因子与其他海拔来源的白桦幼苗差异显著(P<0.05);经过一个生长季后(T2),不同海拔来源的白桦移栽幼苗与海拔750 m野生白桦幼苗(CK1)之间各项生理因子差异性变得不显著(P>0.05),而与海拔1200 m和1400 m野生白桦幼苗之间的差异性仍显著(P<0.05)。
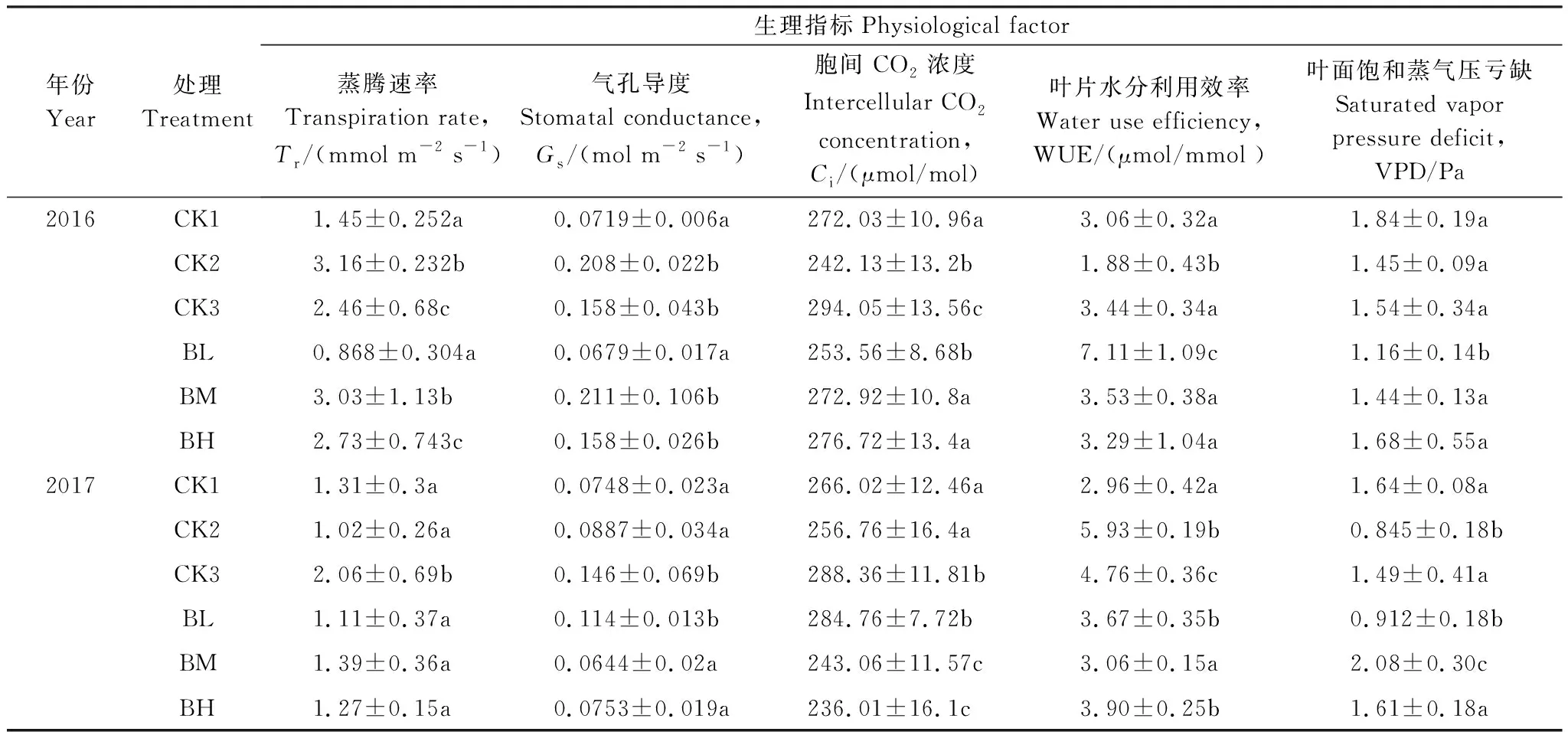
表4 不同海拔来源白桦幼苗2016年和2017年生理指标
2.3 不同海拔来源白桦幼苗反射光谱特性比较
2.3.1移栽初期(T1)和经过一个生长季后(T2)6种白桦幼苗的光谱反射率曲线
图4是移栽初期(T1)和经过一个生长季后(T2)6种白桦的光谱反射曲线,分别在波长500 nm和780 nm左右的位置有两个低反射区,这是蓝、红光波段的光辐射被叶绿素吸收进行光合作用形成的。移栽初期6种白桦幼苗的光谱反射率较低,在550 nm处有一个反射峰,此处不同海拔来源白桦幼苗(BL、BM、BH)和原海拔白桦幼苗(CK1、CK2、CK3)光谱反射率的差异较其他波段较为明显,大小为:BH>CK3>CK1>BM>BL>CK2;经过一个生长季后,白桦在300—400 nm和1100 nm左右的光谱反射率有所提高,在550 nm反射峰,不同海拔来源白桦幼苗光谱反射率大小为:CK2>BH>CK1>BM>CK3>BL。
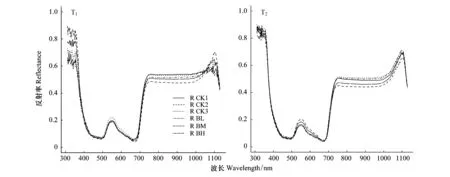
图4 不同海拔来源白桦幼苗2016年移栽初期(T1)和2017年一个生长季后(T2)光谱反射曲线Fig.4 Spectral reflectance curve of Betula platyphylla seedlings under different gradient of light intensity in 2016 (T1) and 2017 (T2)
2.3.2移栽初期(T1)和经过一个生长季后(T2)6种白桦幼苗光谱指数对比
植物光谱指数反应了植物体内部色素含量,包括叶绿素、类胡萝卜素等。由表5可知,移栽初期(T1)不同海拔来源的白桦幼苗之间光谱指数差异较为显著:经过t-检验,除了来源于海拔750 m的移栽苗(BL)与原海拔野生苗(CK1)之间的植被归一化指数(NDVI)、叶绿素归一化指数(ChlNDI)、类胡萝卜指数(CRI)和与结构无关色素指数(SIPI)差异不显著(P>0.05),光化学反射指数(PRI)差异显著(P<0.05),而来源于海拔1200 m和1400 m的移栽白桦苗(BM)和(BH)与来源于海拔750 m的野生白桦苗(BL)之间光谱指数差异显著(P<0.05),但与原海拔野生白桦苗(CK2、CK3)之间光谱指数差异不显著(P>0.05);经过一个生长季后(T2),不同海拔来源的白桦幼苗光谱指数之间差异性均不显著(P>0.05)。

表5 不同海拔来源白桦幼苗2016年和2017年光谱指数
2.3.3 不同海拔来源白桦移栽幼苗与野生白桦幼苗光谱反射率对比
对比不同海拔来源白桦移栽幼苗(BL、BM和BH)与野生白桦幼苗(CK1)之间的光谱比值可得,移栽初期(T1),在可见光范围内,来源于海拔1400 m的白桦幼苗与来源于750 m野生幼苗之间的比值始终高于1,最高可达1.15;来源于海拔1200 m的白桦幼苗与野生幼苗的比值始终小于1,最低值为0.81;来源于750 m的白桦野生幼苗与野生幼苗的比值始终在1左右。经过一个生长季后(T2),不同海拔来源白桦幼苗与野生幼苗的比值保持相同的变化趋势,在波长为400—500 nm之间波长比值始终大于1,在500—700 nm之间比值呈先下降后上升的趋势,在波长600—700 nm之间呈先上升后下降的趋势,来源于海拔1400 m的白桦幼苗比值始终高于其他两个海拔,但总体变化趋势与其他海拔保持相同。
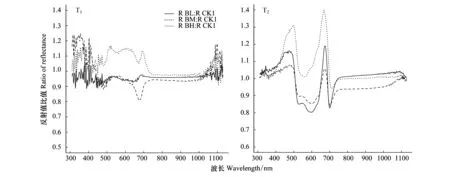
图5 不同海拔来源白桦幼苗2016年移栽初期(T1)和2017年一个生长季后(T2)与野生白桦幼苗光谱反射率对比Fig.5 Comparison of Spectral reflectance between feral Betula platyphylla seedlings and seedlings under different gradient in 2016 (T1) and 2017 (T2)CK1:海拔750 m野生白桦幼苗 Feral Betula platyphylla seedlings from altitude of 750 m;BL:海拔750 m移栽幼苗 Transplanting Betula platyphylla seedlings from altitude of 750 m;BM:海拔 1200 m移栽幼苗Transplanting Betula platyphylla seedlings from altitude of 1200 m;BH:海拔1400 m 移栽幼苗 Transplanting Betula platyphylla seedlings from altitude of 1400 m
2.3.4 不同海拔来源白桦幼苗红边位置比较
红边位置(λRE,red edge)是叶绿素含量较好的评价器,叶绿素会在680 nm周围出现一个荧光峰,因此红边位置是叶绿素含量一个较好的量化指标。通过多重比较分析表明,移栽初期(T1)不同海拔来源白桦幼苗的红边位置存在显著差异(P<0.05),平均值大小为:CK1>CK3>CK2>BL>BM>BH,说明在移栽初期野生白桦的叶绿素含量明显高于移栽的白桦植株;经过一个生长季后(T2),不同海拔来源的白桦幼苗红边位置差异仍显著(P<0.05),但来源于海拔1200 m和1400 m的白桦幼苗间差异不显著(P>0.05),并且来源于海拔750 m的白桦幼苗红边形状表现出一定变化,反射光谱的一阶微分曲线变得较平缓,说明幼苗叶片叶绿素含量下降,与其他海拔叶绿素含量之间的差距变小(图6)。
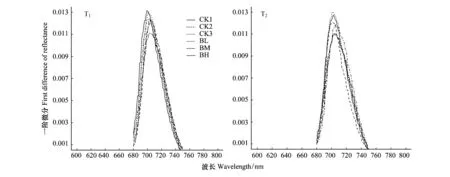
图6 不同海拔来源白桦幼苗2016年移栽初期(T1)和2017年一个生长季后(T2)红边位置Fig.6 Red edge of Betula platyphylla seedlings under different gradient of light intensity in 2016 (T1) and 2017 (T2)
3 讨论
3.1 不同海拔来源白桦幼苗叶片光合特征变化
移栽初期,不同海拔来源的同一物种由于生物的应激性,部分生理性状会发生相应改变,但总体上仍保持原海拔条件下形成的生理性状。与低海拔植物相比,高海拔地区植物叶片的光饱和点较高而光补偿点较低,同时高海拔植物的最大净光合速率要高于低海拔植物[20- 21]。范秀华等[22]对长白山不同海拔梯度的岳桦光响应曲线进行拟合发现,海拔2000 m的岳桦最大净光合速率高于海拔1750 m的岳桦。本研究中,移栽初期(T1)高海拔来源的白桦幼苗光饱和点(LSP)和最大净光合速率(Amax)高于低海拔来源的幼苗,光补偿点(LCP)低于低海拔来源的幼苗。而经过一个生长季后,不同海拔来源的白桦幼苗最大净光合速率(Amax)和光饱和点(LSP)差异性变得不显著(P>0.05)。有研究表明,同一物种长期在相同环境下,其生理性状具有明显的趋同性。Christian等[23]发现非本地的红松与本地的同种红松移栽到相同环境条件下具有与本地种极其相似的生理性状。光饱和点(LSP)、光补偿点(LCP)和最大净光合速率(Amax)等反应了植物对光的应用能力,说明不同海拔来源白桦幼苗的光合能力具有极大的相似性。
同时有研究发现,高海拔地区的植物具有较低的水分利用率(WUE)和较高的蒸腾速率(Tr)[24- 26]。本研究中移栽初期不同海拔来源的白桦幼苗符合上述结论,但在经过一年的适应后,不同海拔的移栽幼苗(BH、BM、BL)与海拔750 m野生幼苗(CK1)蒸腾速率的差异性不显著(P>0.05),水分利用率也呈现极大的相似性,说明不同海拔来源的白桦幼苗在相同环境下,本身的生理性状发生改变。
3.2 不同海拔来源白桦幼苗叶片光谱变化
植物叶片反射光谱指数与叶片色素含量有关,植物叶片光合色素主要包含叶绿素及类胡萝卜素,其中NDVI、ChlNDI等指数与叶绿素含量成正比,而SIPI、CAI等指数与类胡萝卜素含量成正比。对比T1和T2两个时期可以发现,移栽初期不同海拔来源的白桦幼苗各项光谱指数差异性显著(P<0.05)。与野生白桦(CK1)进行对比发现,来源于海拔750 m的白桦幼苗(BL)与野生白桦光谱指数差异不显著(P>0.05),主要原因是两种白桦海拔来源相同,生境相似;而其他海拔来源的白桦幼苗由于海拔梯度造成的环境差异,导致光谱指数差异显著(P<0.05)。有研究表明,叶片叶绿素含量也随着海拔的升高发生变化[27- 28],卢文敏等[29]发现不同海拔间岳桦叶片光谱反射率及光谱指数有较大差异。而在T2时期移栽幼苗之间上述指数之间无显著差异(P>0.05),并与环境中原生幼苗(CK1)相比无显著差异(P>0.05),说明环境变化导致移栽幼苗叶绿素含量发生变化。同时红边位置λRE与叶绿素荧光有很强的相关性。叶绿素含量越高,红光区域吸收增加,导致红边向长波移动[30]。本研究中对比T1和T2两个时期可以发现,不同海拔来源移栽幼苗的红边位置λRE明显发生改变,进一步证明不同海拔来源的白桦幼苗叶绿素含量发生变化。
3.3 不同海拔来源白桦幼苗叶片光谱指数与光合能力的关系
植物叶片反射光谱指数与光合能力也密切相关。其中叶片PRI指数对环境变化极为敏感,是研究植物叶片光合能力的常用指标,与叶片的Pn成正相关关系[31- 32]。本研究中在移栽初期不同海拔的移栽幼苗(BH、BM、BL)与原海拔野生幼苗(CK1、CK2、CK3)之间PRI差异显著(P<0.05),同时不同幼苗的光合能力Pn之间也差异显著(P<0.05)。经过一个生长季之后移栽幼苗(BH、BM、BL)与海拔750 m野生幼苗(CK1)之间叶片PRI与光合能力Pn之间差异不显著(P>0.05),但与海拔1200 m和1400 m野生幼苗(CK2、CK3)之间呈显著差异(P<0.05)。PRI与类胡萝卜素或叶绿素含量成负相关[33],说明移栽幼苗的色素含量发生变化,并与环境中原生幼苗的色素含量无显著差异(P>0.05)。
总之,由于海拔梯度带来的环境差异,导致不同海拔原生幼苗之间存在生理性状上的差异,本研究发现,将不同海拔来源的白桦幼苗迁入新环境后,迁入早期各个海拔来源的幼苗仍保留原海拔条件下形成的生理特性,并与新环境中原生幼苗在生理性状上存在显著差异(P<0.05)。研究还表明,在经过较长时间的环境驯化后,不同海拔来源的白桦幼苗与新环境中野生幼苗生理性状之间的差异性变得不显著(P>0.05),说明白桦具有良好的环境适应性,能够通过调整自身的生理性状适应新环境,这也进一步揭示了白桦作为先锋种植物在演替初期的重要作用。本研究主要在低海拔条件下进行,至于白桦在其他海拔梯度上的适应能力,以及在更长期的环境适应后其生理性状的变化有待进一步研究。
