鄱阳湖湿地灰化苔草种群生产力特征及其水文响应
2018-08-29周云凯白秀玲宁立新
周云凯,白秀玲 ,*,宁立新
1 河南大学环境与规划学院,开封 475004 2 北京师范大学地理科学学部,北京 100875
植物是湿地生态系统中重要的组成部分之一,也是主要的初级生产者,其空间分布及生长发育受气候、水文、土壤以及地形等多因素影响[1- 2]。大量研究表明,湿地植物往往沿某个环境梯度(如水分、盐分、高程、温度等)呈较为明显的带状或环带状分布[3- 6]。对于淡水湖泊湿地而言,水分条件是影响湿地植物的关键因素[7- 9],湿地水分条件(淹没深度、淹没频率等)从湖岸到湖心方向随高程降低表现出一定的带状变化特点,形成不同植物的适生环境,从而引起植物的空间分布也呈现出相应的带状分异,这是湿地植物对水分条件差异性适应和响应的结果[10-12],每种湿地植物都占据着一定的水分生态位[13]。而在同一植物分布带中,受水分梯度和水位波动的影响,植物的生长发育过程不尽相同,植物会对不同的水分条件产生趋异反应,表现出不同的生态和生理特征[14-17]。
长期以来,湿地植物与水之间的作用关系一直都是湿地生态学研究的重要领域。其中,湿地水分条件如何影响植物生长与生存、湿地水情变化如何影响植物的空间分布与群落演替又是湿地生态学研究的热点问题[18- 21]。针对湿地植物生长发育与水分条件之间关系,许多学者已开展较为广泛的研究。如王丽等[17]研究了三江平原小叶章(Calamagrostisangustifolia)生物量积累及其分配对不同水分梯度的响应;栾金花等[22]研究了三江平原不同水分梯度下漂筏苔草(Carexpseudocuraica)株高的生长变化规律;李亚芳等[23]则研究了洞庭湖湿地不同高程短尖苔草(Carexbrevicuspis)生长繁殖策略与水位变化之间的关系。然而当前相关研究更多侧重于场地模拟控制试验,而对野外实地监测研究相对较少,通过人为设定或控制的水分梯度,往往难以反映湿地复杂的水文条件和特定植物的水文经历,而开展自然状态下湿地植物生长发育过程及其与水分条件之间关系研究,对于深入了解湿地植被格局形成、植物群落演替及进行湿地生态调控与恢复具有更为重要的理论和现实意义。
鄱阳湖湿地是我国重要的淡水湖泊湿地之一,受长江及“五河”(赣江、抚河、饶河、信江和修水)来水的共同作用,水位季节变化明显,呈现丰水期和枯水期交替出现的独特水文节律[24],湿地洲滩各高程带“淹没-出露”过程不一,这种不同的水文经历使洲滩植被沿高程梯度呈现出较为明显的带状分布格局[13]。近年来,随着气候变化和三峡水利工程兴建运行等人类活动的干扰,鄱阳湖水文情势发生显著改变,主要表现为丰水期水位降低、枯水期提前和低水位持续时间延长[25- 26],这势必会影响到湿地植物的生长发育、分布格局与群落结构,进而影响湿地生态系统功能的发挥[12,27]。但目前针对鄱阳湖湿地植物与水文变化之间关系研究还十分薄弱,对于湿地典型植物在生长发育过程中对不同水文条件采取的适应策略以及植物定植生长所需的水文条件阈值尚不清晰,为此,本文选取鄱阳湖湿地洲滩优势植物——灰化苔草作为研究对象,基于野外实地观测,研究自然状态下鄱阳湖湿地不同高程带灰化苔草种群发育过程及其与水文条件之间的响应关系,以认清鄱阳湖湿地节律性水文变化背景下灰化苔草的生态适应策略、响应方式和水文生态阈值,为进一步揭示鄱阳湖湿地植被带状分布格局形成及植物群落演替机制提供基础理论支持,同时也为鄱阳湖湿地水文调控、生态恢复与生物多样性保护提供重要科学依据。
1 材料与方法
1.1 研究区概况
鄱阳湖位于江西省北部,长江中下游南岸,地理位置24°29′14″—30°04′41″N、113°34′36″—118°28′58″E。上承“五河”(赣江、抚河、饶河、修水、信江)及区间来水,经由湖口调蓄后汇入长江,与长江存在着复杂的水文、水动力交互,是一个典型的吞吐型湖泊。由于受上游来水和长江的顶托、拉空和倒灌等作用的综合影响,鄱阳湖水位年内变化剧烈,年内最大变幅可达9.7—15.79 m,呈现出“洪水一片,枯水一线;高水是湖,低水似河”的独特自然景观[6,28]。湖区地处丘陵平原区,四周群山环绕,整个湖盆自东南向西北倾斜。以松门山为界,可将鄱阳湖分为南北两部分,南部宽浅,为主湖区;北部狭深,为入江水道。湖泊地貌主要有洲滩、岛屿、内湖、水道和汊港等类型[29]。该地区气候上属于亚热带季风气候,雨热同期,四季分明,年均气温16.5—17.8℃,无霜期250—280 d,年均日照时数约为1970 h,年均太阳总辐射量可达4500×106J/m2,而多年平均降水量为1350—1700 mm[30]。湖区洲滩土壤主要有草甸土、草甸沼泽土和沼泽土等类型[31]。由于水热资源较为充足和水位周期性涨落变化,鄱阳湖区发育了大片的天然湿地,成为我国生物多样性最为丰富的地区之一[32]。从湿地边缘向中心,植物呈较为明显的带状或环带状分布特征,依次分布着以狗牙根(Cynodondactylon)、蒿(Artemisiaspp.)等为主的中生植物群落,以芦苇(Phragmitesaustralis)、南荻(Triarrhenalutarioriparia)等为主的挺水植物群落,以苔草(Carex)、虉草(Phalarisarundinacea)等为主的湿生植物群落和以马来眼子菜(Potamogetonmalaianus)、苦草(Vallisnerianatans)、菱(Trapabispinosa)、荇菜(Nymphoidespeltata)等为主的沉水/浮叶植物群落[6,33]。其中灰化苔草是鄱阳湖湿地分布最广的洲滩植物种,主要分布在洲滩的中低高程带上。
1.2 样地设置
本研究选取灰化苔草分布集中的蚌湖东侧修水入湖口洲滩作为野外观测样地,该洲滩高程在15 m左右,洪水期被湖水淹没,枯水期出露,洲滩上植物沿高程呈现出较为明显的带状分布,随着高程由低到高依次分布着虉草、灰化苔草、南荻等植物群落,其中灰化苔草面积最大,生长也较为茂盛,分布在12—14.5 m的高程带上。洲滩未经开垦、放牧、火烧等人为干扰,自然环境保存良好。分别在洲滩四周按与湖岸垂直方向(沿高程或水分梯度)各布设1条样带,样带宽度为20 m,样带长度以整个洲滩上灰化苔草出现的最低和最高高程来进行确定;在每个样带内按0.5 m高程距离布设采样点,在同一高程带上,按等距离设立3个面积为1 m×1 m的样方,用GPS-RTK(南方测绘,灵锐S86)确定各样方的高程和地理位置。
1.3 研究方法
2013年3—5月在设置的样地内进行灰化苔草种群生态观测和样品采集(灰化苔草在2月下旬萌发,3—5月为其生长季,5月下旬洲滩被湖水淹没,灰化苔草死亡或沉入水底休眠,完成其一个生长周期),采样时间间隔15天左右。对每个样方分别测量灰化苔草种群的高度、密度、地上生物量和地下生物量等生态指标。其中,用标尺测定灰化苔草株高;用计数法测定种群密度;地上生物量采用收获法,地下生物量采用挖掘法。采集10株植物样品带回实验室处理,先用清水洗净地下根系,用吸水纸吸干后,分别对地上和地下部分进行湿重称量,之后在鼓风干燥箱中先于105℃下杀青10分钟,再在80℃下烘干至恒重,称其干重,最后换算成单位面积的地上及地下生物量,两者相加计算出总生物量。用总生物量干鲜比(DW/FW)反映植物体含水量的多少(干鲜比=干重(Dry Weight)/湿重(Fresh Weight) ×100%);用生物量增长速率分析方法中的绝对增长速率(Absolute growth rate, AGR)和相对增长速率(Relative growth rate, RGR)反映灰化苔草种群生物量的积累状况,计算公式分别为:
(1)
(2)
式中Bi、Bi+1分别是ti、ti+1时刻的灰化苔草种群的生物量(g/m2)。
1.4 数据处理与分析
借助Excel 2007进行数据处理和制图。利用SPSS 19.0对测定数据进行统计分析,采用相关分析确定灰化苔草种群生态指标与水文参数之间的相关性;通过单因素方差分析(one-way ANOVA)和LSD多重比较法进行不同高程带生态指标差异显著性检验。
2 结果与分析
2.1 灰化苔草种群特征及其变化
由于13 m以下高程带的灰化苔草在3月下旬即被湖水淹没,因此,在进行灰化苔草种群发育过程研究时,主要针对13 m及以上高程带的灰化苔草种群进行分析。
2.1.1 密度与株高
初步研究表明,在生长季内,各高程带灰化苔草的种群密度变化趋势基本一致,表现为随时间推移而逐渐增加,但变化过程略有差异(图1)。14.5 m高程带灰化苔草种群密度在3月下旬之前增加较快,而之后基本保持稳定,维持在1875—1900株/m2;13 m和13.5 m高程带的种群密度在3月上旬至4月中旬增加较为缓慢,4月下旬之后增加速度较前期有所加快;而14 m高程带种群密度在3月下旬之前增加较慢,之后开始加快,到4月下旬增加速度再次放缓。各高程带灰化苔草种群的平均株高与密度变化趋势基本一致(图1),整体表现为随时间推移而逐渐增高,但不同生长阶段增长幅度存在明显差异。在4月下旬之前,株高的增长速度随高程的增加而增大,而4月下旬之后,14 m和14.5 m高程带增长速度明显减慢,而13 m和13.5 m高程带增长速度则快于前期。

图1 灰化苔草种群密度与株高变化Fig.1 Changes of height and density of Carex cinerascens population in different elevation zones
2.1.2 生物量
各高程带的灰化苔草生物量变化及累积过程有所不同(图2)。在生长季内地下生物量与总生物量一直处于增加状态;而地上生物量除13 m高程带呈不断增加外,其他高程带上均表现为先增加后减少的变化过程。从绝对增长速率来看,各高程带灰化苔草地下生物量最大绝对增长速率出现在4月下旬—5月上旬(图3),而地上生物量与总生物量的最大绝对增长速率出现时段不同,整体表现为13.5 m及以上高程带出现在4月中旬,13 m高程带则出现在4月下旬—5月上旬;这表明灰化苔草快速生长期出现时段随高程而变化,高程越高,则出现越早。从相对增长速率来看,各高程带地下生物量与总生物量一直处于累积过程,地上生物量在13 m高程带上也一直处于累积过程,而13 m高程带以上则在4月下旬之前处于累积过程,之后开始衰退,生物量出现降低。

图2 灰化苔草种群生物量变化Fig.2 Changes of aboveground, underground and total biomasses of Carex cinerascens population in different elevation zones
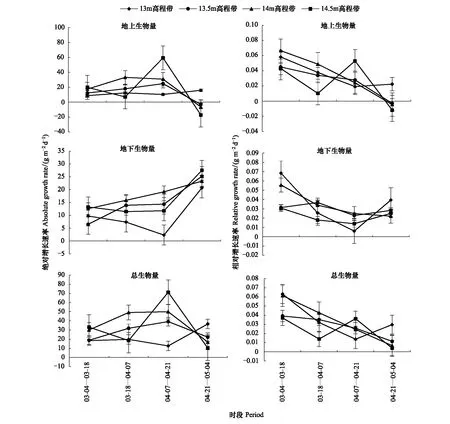
图3 不同生长阶段灰化苔草种群生物量增长速率变化Fig.3 Changes of absolute growth rate (AGR) and relative growth rate (RGR) of aboveground, underground and total biomasses of Carex cinerascens population in different elevation zones
2.1.3 干鲜比
各高程带灰化苔草在生长季内的干鲜比随时间变化过程基本一致(图4),整体均呈增加趋势,在3月上旬灰化苔草生长的初期阶段,干鲜比较低,在20.8%—23.2%之间;进入4月中旬的快速生长期阶段,灰化苔草的干鲜比有所增大,为23.3%—26.1%;到5月上旬的成熟期,灰化苔草干鲜比达到整个生长季的最大值,为26.8%—29.1%。从灰化苔草干鲜比的变化过程可以看出,生长初期的干鲜比要小于生长后期,这表明随着时间的推移,灰化苔草植物体内的水分含量逐渐减少,而积累的干物质含量逐渐增多。

图4 灰化苔草干鲜比变化 Fig.4 Change of DW/FW of Carex cinerascens population in different elevation zones
2.2 鄱阳湖湿地水文条件空间分异
淡水湿地植物的分布和生长发育是对周期性水文节律长期适应的结果。在灰化苔草相邻的两个同期生长季间(2012年5月4日—2013年5月4日),鄱阳湖水位(星子站,吴淞高程,下同)变化十分显著(图5),2月份湖水位达到最低,为8.91 m(2013年2月6日),8月份湖水位最高,为19.64 m(2012年8月13日),最大水位差达10.73 m。在最高与最低水位之间,湖泊水位呈现出明显的涨水期和退水期。对于鄱阳湖洲滩不同高程带而言,由于水文经历的差异,在各高程带上形成了不同的水文条件,表现出一定的水分梯度变化。在此将各高程上的水文条件以淹没期平均水深(淹没的总水深与淹没天数的比值)、淹没频率(淹没天数与计算总天数的比值)和平均距湖水面高度(高程与日水位差值之和与计算总天数的比值,水面以下为-,水面以上为+)来进行反映。在研究期间,洲滩上灰化苔草分布的下限12 m高程带大部分时间被湖水所淹没,淹没期水深在0.02—7.64 m之间,平均值为3.89 m,相邻同期生长季内淹没频率为60.38%,平均距湖水面高度为-1.73 m;随着高程的增加,洲滩淹没天数逐渐减少,而出露天数逐渐增多,至洲滩灰化苔草分布上限14.5 m高程带上,淹没期平均水深减小到2.78 m,该高程带上淹没频率降低到42.08%,平均距湖水面高度增大为0.77 m(表1)。总体来看,洲滩上灰化苔草定植与生长的水文条件阈值为淹没期平均水深2.78—3.93 m,淹没频率42.10%—60.38%,而平均距湖水面高度则为-1.73—0.77 m。
2.3 不同高程灰化苔草种群特征差异及对水文条件响应
选取灰化苔草接近成熟期的4月下旬进行不同高程带灰化苔草种群特征的差异分析。由表2可知,灰化苔草种群密度随高程呈波动变化,差异性分析显示不同高程带灰化苔草种群密度差异不显著(F=2.171,P>0.05),这说明高程形成的水分梯度对种群密度无明显影响。
不同高程带灰化苔草平均株高随着高程的升高而逐渐增加,高程间存在显著差异(F=5.879,P<0.05),14.5 m高程的平均株高达到117.5 cm,是13 m高程的两倍。多重差异性比较发现,13 m高程带灰化苔草株高与13.5 m高程带差异不明显,而与14 m及14.5 m高程带形成显著差异,其他高程带间差异则不明显,这说明13.5以下高程的水文条件对灰化苔草株高的增加起到抑制作用,而13.5 m及以上高程带的水文条件更有利于灰化苔草的伸长生长。
不同高程带灰化苔草生物量均随高程的升高而增大,且高低滩位间相差一倍以上,高程间亦存在显著差异(地上生物量:F=5.022,P<0.05;地下生物量:F=5.485,P<0.05;总生物量:F=6.599,P<0.05)。多重差异性分析显示,13 m高程带的地下生物量与总生物量与其他高程带存在显著差异,其他高程带之间则差异不显著;而地上生物量13 m高程与13.5 m高程无明显差异,但与14、14.5 m高程带差异显著,而其他高程带间的地上生物量无明显差异。可以看出,13.5 m及以上高程带的水文条件更有利于灰化苔草生物量的形成与积累,而13 m高程带的水文条件对生物量积累相对不利。
将4月下旬灰化苔草各生态指标与淹没期平均水深、淹没频率、平均距湖水面高度3个水文参数分别做相关分析,结果显示,除密度以外,其他各指标与水文参数均呈显著负相关(P<0.01),这进一步说明水文条件对灰化苔草生长发育产生重要影响。
由上述分析可知,灰化苔草生长发育对高程形成的不同水分梯度产生差异性响应。在13.5 m以下的低滩位,由于频繁的过度淹没而使灰化苔草生长受到水分条件的抑制,植株矮小,生物量积累相对较少;而在13.5 m及以上中高滩位,形成灰化苔草生存有利的水文环境,更利于灰化苔草的伸长生长和生物量的形成与累积。结合前面水文参数的分析,可对春季生长季内灰化苔草生长发育的适宜水文条件做出初步判断,有利于灰化苔草生长所需的淹没期平均水深阈值为 3.70—2.78 m,淹没频率阈值为42.08%—43.17%,平均距湖水面高度阈值为-0.23—0.77 m。

图5 鄱阳湖湿地星子站水位变化Fig.5 Water-level variation of Xingzi hydrological station in Poyang Lake wetland during 2012/5/4—2013/5/4

高程带Elevation/m淹没期水深Water depth of inundation period/m范围Range平均值Average淹没频率Inundation frequency/%平均距湖水面高度Average height from water surface/m120.02—7.643.8960.38-1.7312.50.03—7.143.9352.73-1.23130.02—6.643.9246.45-0.7313.50.14—6.143.7043.17-0.23140.08—5.643.2442.620.2714.50.05—5.142.7842.080.77
3 讨论
3.1 灰化苔草水分适应策略
淡水湿地植物的空间分布及其生长发育过程与高程、水文状况、土壤性质以及植物种类等因素密切相关,高程及其形成的水文条件起着至关重要的作用[2,34]。同一植物种类处在不同的水文条件下,植物会通过形态和自身生理机能的调整来适应变化的水文环境,形成能够反映生境条件的生态与生理特征[35]。对于水位周期性波动的淡水湿地来说,洲滩或湖(河)滨带经常会遭受不同程度的淹水胁迫[26],湿地植物往往通过枝条伸长生长的逃避策略或者休眠、缓慢生长的耐受策略来应对水分胁迫[36-37],不同的淹水条件下各种植物会采取不同的适应策略。已有研究表明,在淹水条件下双穗雀稗(Paspalumdistichum)、空心莲子草(Alternantheraphiloxeroides)和牛鞭草(Hemarthriaaltissima)等湿地植物会通过伸长生长来逃离淹水环境[38],而虉草和狗牙根(Cynodondactylon)则通过减缓茎的伸长和不产生新的分枝等方式来提高自身对水淹的耐受性[39]。

表2 不同高程带灰化苔草种群特征及差异(n=13)
数据为平均值±标准差;每列数据右上角标有不同字母表示高程间存在显著差异(P<0.05)
本研究发现,灰化苔草在生长季会根据水淹状况来调整自身的生存策略。13 m以下高程带的灰化苔草在3月下旬即被湖水淹没,长期的水淹导致水中供氧和光照不足,光合作用和叶绿素合成受到严重抑制,灰化苔草通过地上植株死亡和地下根系休眠的耐受策略来增加存活率;13 m高程带在4月上旬一直处于半水淹状态,该期间灰化苔草株高和生物量增长速度减缓,而在4月中下旬湖水位发生波动式的下降,灰化苔草经历间歇性的半水淹,但大部分时间植株完全出露水面,这一期间灰化苔草生长发育过程明显加快,株高及生物量的增加幅度达到整个生长季的最大值,灰化苔草通过快速伸长生长的逃避策略来应对短期的间歇性半水淹环境;在13.5 m及以上高程带上,灰化苔草5月上旬之前基本未遭受淹水胁迫,株高和生物量在各生长阶段均高于13 m高程的低滩位,且生长后期差异更加明显,然13.5 m及以上高程带由于萌芽较早,在4月下旬即步入成熟期,该阶段虽然株高仍在增长,但生物量开始逐渐向地下部分转移以完成生活史,导致地上生物量出现一定程度的降低,但13.5 m和14 m高程带的总生物量仍在增加,而14.5 m高程带由于提前进入枯萎衰落期,有机质合成减弱,呼吸消耗增多,总生物量有所下降。在5月中旬之后,湖水位大幅上涨,鄱阳湖进入丰水期,灰化苔草分布洲滩完全被湖水淹没,各高程带地上部分逐渐死亡,灰化苔草进入休眠状态,完成其完整的春季生长季。由此可见,各高程带灰化苔草对不同的水文经历采取差异性适应策略,这种生存策略是灰化苔草对鄱阳湖湿地周期性水文节律长期适应进化的结果。
3.2 灰化苔草水分利用策略
湿地植物在不同生长阶段会根据实际的水分条件来调节自身的水分利用策略,通过植物干鲜比可以对其采取的水分利用策略予以反映。植物干鲜比标志着植物体含水量的多少,它是植物对水分条件以及植物不同生长阶段所表现出的一种响应,湿地植物对不同的水分条件会产生趋异响应[17]。上官铁梁等[40]通过对汾河河漫滩湿地植物群落研究发现,植物干鲜比随着群落水分增加而增大;而王丽等[17]对三江平原湿地小叶章群落研究也发现,在生长季初期小叶章主要通过储水策略来保持体内一定的储水量,干鲜比较低,在生长季后期由于可获得足够的水分资源,干物质积累多且迅速,干鲜比较高。在本研究中,鄱阳湖湿地灰化苔草也采用相似的水分利用策略,在生长初期干鲜比较低,而生长中后期则显著增大,这种变化一方面与灰化苔草自身的生长特性有关,另一方面也与鄱阳湖湿地特定的水文过程有关。在灰化苔草生长初期,由于植株矮小,气温较低,灰化苔草蒸腾作用和呼吸作用较弱,而此时正值鄱阳湖区的少雨季,湿地处于一年当中的低水位期,水分条件十分有限,灰化苔草主要采取储水策略以维持自身生长对水分的需求,干鲜比较低;到灰化苔草生长的中后期,由于降水增多以及湖水位的抬升,水分资源较为充足,灰化苔草进入快速生长期,但该时期灰化苔草的蒸腾作用和呼吸作用明显增强,植株体内的水分被大量消耗,而由光合作用形成的干物质较前期增多,引起干鲜比出现增大。灰化苔草生长过程中干鲜比的变化一定程度上也反映出其对鄱阳湖湿地水位节律性变化的生态适应。
3.3 灰化苔草水文条件阈值
每种湿地植物在生长发育过程中对水文条件都有特定的需求,在某个阈值内植物可以存活并生长。众多水文因子中,水深和淹没频率是影响湿地植物定植生长、繁殖和分布的重要环境因子[41- 42]。对于湿地沉水植物而言,其整个生长发育过程均在水中进行,水深通过影响水下光照、风浪和底泥特征来作用于植物生长繁殖,如黑藻(Hydrillaverticillata)和苦草(Vallisnerianatans)分别在水深0.5—2.5 m和0.5—5.6 m才能够定植生存,当水深继续增加时,光照强度会低于光补偿点,光合作用受到抑制,植物将难以存活[43-44]。而对于某些湿地挺水植物和湿生植物来说,在生长过程中会经历一定程度的水淹和干旱,因此,除水深因子外,植物定植生长也会受到淹没频率的影响[41]。已有研究表明,适度的干湿交替有利于挺水植物和湿生植物的生长繁殖,而长期的过度水淹和无水淹会减弱植物光合作用和养分吸收能力,对植物生长起到明显的抑制作用[35,45]。但不同湿地植物对水文条件的喜好性和耐受性有所差异,如鄱阳湖湿地南荻生长于相对干旱的环境中,不能忍受长时间的洪水淹没,其主要集中分布于平均水深2.2—2.4 m、淹没频率15%—40%的区域,而假俭草(Eremochloaophiuroides)对水文条件适应范围较宽,在平均水深1.8—14.6 m、淹没频率12%—100%的区域均可定植生长[42]。灰化苔草是鄱阳湖湿地洲滩典型的湿生植物,生长于相对湿润的环境中,对水淹和干旱有一定的耐受能力,具有较大的水分生态幅[46],适度的水淹和露干交替有利于生长所需水分的获取和光合作用的进行,可促进灰化苔草的伸长生长和生物量的累积,而长期水淹或干旱将超出灰化苔草的耐受极限,不利于植株体的存活和种群的发育。本研究发现,灰化苔草在淹没期平均水深2.78—3.93 m、淹没频率42.1%—60.38%、平均距湖水面高度-1.73—0.77 m的水文条件下能够定植生长,而淹没期平均水深3.70—2.78 m、淹没频率42.08% —43.17%、平均距湖水面高度-0.23—0.77 m的水文环境则对灰化苔草的生长更为有利,其种群的平均高度和生物量均达到最大。
许多研究表明,部分湿地植物种群的生态特征(如株高、生物量)对水文因子(如水深)的响应符合高斯分布[47-49],然在野外环境中,影响湿地植物种群的环境因子众多,除水文条件以外,植物种内及种间竞争、土壤养分、温度、降水等环境因子也会产生影响,实际上湿地植物种群生态特征是多环境因子综合作用的结果[50],鄱阳湖湿地灰化苔草种群生态指标沿水分梯度并未呈现出明显的高斯分布,而是在水分阈值内大致随水分梯度的降低而增加,这种分布格局的形成与其他环境因子的空间分异及作用强度也存在一定关系,但对于其具体的形成机制和作用机理还有待今后进一步地深入研究。
4 结论
(1)鄱阳湖湿地灰化苔草作为洲滩优势植物种群之一,在春草生长季内各高程带种群变化规律基本一致,但由于高程带间水文条件与水文经历的不同,灰化苔草生长发育过程对节律性水文波动产生差异性适应和响应。在13.5 m以下的低滩位,由于频繁的水淹而使灰化苔草生长发育受到水分条件的抑制,植株矮小,生物量积累相对较少;而在13.5 m及以上中高滩位,形成灰化苔草适生的水文环境,更利于灰化苔草的伸长生长和生物量的形成与积累。
(2)灰化苔草在不同生长阶段会根据实际的水分条件来调节自身的水分利用策略。生长初期灰化苔草主要通过储水策略来应对水分相对缺乏形成的干旱胁迫;而生长末期则主要通过增加干物质含量来应对湖水位上涨形成的淹水胁迫。
(3)灰化苔草定植与生长发育对水文条件存在一定的适应范围。在淹没期平均水深2.78—3.93 m、淹没频率42.1%—60.38%、平均距湖水面高度-1.73—0.77 m的条件下灰化苔草能够定植生长;而淹没期平均水深3.70—2.78 m、淹没频率42.08%—43.17%、平均距湖水面高度-0.23—0.77 m的水文环境则更有利于灰化苔草的生长发育。
