花生甘油-3-磷酸酰基转移酶(GPAT)基因的克隆及表达分析
2018-08-29郝翠翠梁成伟李昊远陈明娜潘丽娟禹山林迟晓元
郝翠翠,梁成伟,石 蕾,李昊远,3,陈明娜,潘丽娟, 王 通,王 冕,禹山林,陈 娜*,迟晓元*
(1.青岛科技大学海洋科学与生物工程学院,山东 青岛 266042;2.山东省花生研究所,山东 青岛 266100;3.哈尔滨工业大学(威海)海洋科学与技术学院,山东 威海 264209)
花生(ArachishypogaeaL.)是我国重要的油料作物、也是一种经济型农作物及出口创汇作物。我国花生产量约占国内油料作物总产量的50%[1-2]。我国每年有一半以上的花生被用来榨油,花生油消费量在每年2000 kt以上,占食用植物油消费量的25%左右。目前,我国食用植物油的需求呈现供不应求的局面。开展油料作物研究,提升植物油自给率迫在眉睫。花生在保障我国食用植物油供给中具有明显的潜力和优势,研究花生油脂合成过程及机理,提高花生油脂含量,才能符合未来对于植物油脂的消费需求。
GPATs在植物油脂生物合成过程中扮演着重要角色,其主要作用是催化酰基辅酶A(acyl-CoA)上的脂肪酸连接到3-磷酸甘油的sn-1位点上,形成溶血磷脂酸(Lysophosphatidic acid,LPA),是生物合成过程中的第一步反应[3]。GPAT基因已经在拟南芥(Arabidopsisthaliana)、水稻(Oryzasativa)、向日葵(Helianthusannuus)、菠菜(Spinaciaoleracea)、油桐(Jatrophacurcas)、番茄(Lycopersicumesculentum)和豌豆(Pisumsativum)等多种植物中克隆出来[4]。其中,模式生物拟南芥GPAT基因家族有10个成员:ATSl、GPAT1-9。
已有研究发现,拟南芥AtGPAT9基因与种子含油量有关,可能参与了TAG的合成,同时,其他植物中的GPAT在提高种子含油量和油脂质量方面,也具有非常重要的作用[5-7]。为探究GPAT蛋白在花生油脂生物合成和逆境胁迫中的重要作用,本研究从课题组已有的cDNA文库中获得2个新的酰基载体蛋白的基因全长,分别命名为AhGPAT3和AhGPAT5。将得到的完整序列进行生物信息学分析,同时测定这两个基因在花生不同组织器官、种子发育不同时期、多种逆境胁迫及激素处理下的表达特性,为进一步研究花生GPAT在调控油脂生物合成中的功能奠定了理论基础。两基因的GenBank注册号分别为KP109922和KP109923。
1 材料与方法
1.1 植物材料
供试材料为花育33,由山东省花生研究所提供。播种花生种子,置于光照培养箱中培养,光周期16 h光照/8 h黑暗,温度28℃/22℃。在花生生长到三叶期时,分别采集根、茎、叶、子叶和下胚轴,盛花期采集花生花。花生开花下针后,分别采集其下针后12、24、36、48和60 d的种子。
花生生长到三叶期后进行多种非生物胁迫和激素处理。盐(NaCl 200 mmol/L)、低温(4℃)、干旱(15% PEG6000)均处理0、4、8、12、24、72 h。水杨酸(SA,2 mmol/L)、过氧化氢(H2O2,10 mmol/L)均处理0、4、8、12、24、36、72 h;赤霉素(GA,100μmol/L)、乙烯利(ET,5 mmol/L)和脱落酸(ABA,100 μmol/L)均处理0、8、12、24、48、72 h;茉莉酸(JA,100 μmol/L)处理0、4、8、12、24、72 h。对照组:ABA和GA以相同浓度乙醇作为对照,其他以清水作对照。除低温处理外,在进行处理时,应先将花生三叶期幼苗从土中小心取出,清洗去除根部泥土,再置于相应条件下接受处理。除低温、盐和干旱处理外,其余处理还需要用溶液涂抹花生叶片。在处理的不同时间段分别取花生幼苗的叶片,放入液氮中冷冻保存备用。
1.2 总RNA提取与cDNA合成、目的基因的扩增
在NCBI上搜索到拟南芥两个蛋白序列,分别是AtGPAT3(NP_192104)和AtGPAT5(NP_187750)。利用BioEdit软件从花生cDNA文库(共计36741条EST序列)中搜索与上述两个拟南芥的氨基酸序列相似性较高的序列,作为候选基因。最终从已知的花生cDNA文库中找到了2个可能编码甘油-3-磷酸酰基转移酶的基因的全长序列:AhGPAT3和AhGPAT5。根据此序列,使用Primer 5软件设计基因全长引物。引物序列为AhGPAT3-F:5'-ATGGCTAAAATGTTCACCAT-3'和AhGPAT3-R:5'-CTACTTGTTGGAACTGCTGG-3';AhGPAT5-F:5'-ATGGAGTGTGTTGTGTCTGA-3'和AhGPAT5-R:5'-TTACTGTAAAAACGGTTTAA-3'。参照陈娜等[8-9]的方法提取花生叶片总RNA并进行反转录。
以cDNA为模板,通过PCR技术扩增获得目的基因。PCR反应条件如下:94℃ 2min;94℃ 30s,68℃ 30 s,72℃ 2 min,35个循环;72℃ 10 min。将PCR所得目的片段切胶回收,链接到T载体后,转入大肠杆菌DHa5中,筛选阳性克隆进行测序。将测序得到的序列进行拼接,得到GPAT基因全长序列,初步分析后提交到GenBank中。
1.3 氨基酸序列分析
采用NCBI网站(http://www.ncbi.nlm.nih.gov)的ORF finder查找基因的开放读码框;使用ProtParam(http://web.expasy.org/protparam/)分析蛋白质的基本物理化学参数;采用TMHMM 2.0 Server分析蛋白跨膜结构;利用PSIPRED(http://bioinf.cs.ucl.ac.uk/psipred/)分析蛋白二级结构;用GSDS(Gene structure display server)对基因的外显子/内含子结构进行分析。
1.4 多序列比对与系统发育分析
利用NCBI网站的BlastP工具分别对克隆得到的两个基因进行基因序列的相似性及同源性查找,并利用这些序列进行基因同源性比较。结合Clustal X和BioEdit软件将下载序列与AhGPAT3和AhGPAT5序列进行比对,通过比对寻找序列之间的相似区域和保守性位点,寻找可能的进化关系。
为了更好地分析确定GPAT基因的进化关系,利用formatdb程序建立一个拥有6个基因组的数据库。这6个基因组分别为:大豆(Glycinemax)基因组,小立碗藓(Physcomitrellapatens)基因组,蒺藜苜蓿 (Medicagotruncatula)基因组,团藻(Volvoxcarteri)基因组,莱茵衣藻(Chlamydomonasreinhardtii)基因组和拟南芥(Arabidopsisthaliana)基因组数据。基因组数据均从phytozome(http://www.phytozome.org/)数据库下载获得。起始源序列用已经通过实验验证的拟南芥GPAT基因,利用BlastP和TBlastN程序,设定E值<1e-50,在新建立的基因组数据库中搜索,然后以新搜索到的基因重复此过程,直到搜索不到新的序列为止[10],最后合并搜索到的所有序列。采用ClustalW软件对合并序列进行多序列比对分析,然后采用MEGA 4.0软件进行系统发生和进化的分析。系统发育树采用Neighbour-Joining(距离邻接法)方法构建。
1.5 荧光定量PCR
采用Roche的LightCycler 2.0 instrument system进行PCR反应。反应方法和条件参照陈娜等[8-9]的方法。采用Actin11为内参基因,3次重复,实验数据使用2-△△Ct方法分析[11]。 荧光定量采用如下引物:
qAhGPAT3-F: 5'-CTACATCACCACCACCAT -3',
qAhGPAT3-R: 5'-GAAGAAGAGAAGAAGAAGAAGAG -3';
qAhGPAT5-F: 5'-GATTCGGATGCTGGACAAG -3',
qAhGPAT5-R: 5'-TCTATATCTGACTCACCAACA -3';
qACT11-F: 5'-TTGGAATGGGTCAGAAGGATGC -3';
qACT11-R: 5'-AGTGGTGCCTCAGTAAGAAGC -3'。
2 结果与分析
2.1 AhGPAT3和AhGPAT5基因的克隆
以花生叶片的cDNA第一条链为模板,进行PCR扩增,链接载体转入大肠杆菌后筛选阳性克隆检测,有特异性条带,表明成功克隆两基因。通过Blast搜索、序列比对和系统发育分析,确定两基因为GPAT家族成员,并且分别与拟南芥AtGPAT3和AtGPAT5基因序列相似性高,聚类在一起,因此,将克隆得到的两基因分别命名为AhGPAT3和AhGPAT5。其中,AhGPAT3基因全长1653 bp,编码550个氨基酸;AhGPAT5基因全长1383 bp,编码460个氨基酸 (图1和图2)。

图2 AhGPAT5基因的核苷酸及氨基酸序列 Fig.2 Nucleotide sequence and deduced amino acid sequence of AhGPAT5
2.2 氨基酸序列分析
对AhGPAT3和AhGPAT5基因编码的蛋白质进行物理化学性质分析,结果显示:AhGPAT3蛋白的理论分子量为62.38 kD,理论等电点为9.02,平均亲水系数0.061。AhGPAT5蛋白的理论分子量为51.42 kD,理论等电点为9.36,平均亲水系数0.223。
AhGPAT3蛋白含的550个氨基酸残基中,Leu(L)含量最高,占氨基酸总数的12.2%;其次为Ser(S)、Val(V)、Phe(F),分别占氨基酸总量的9.5%、8.2%和7.8%。含有55个带负电的氨基酸残基(Asp+Glu),占总数的10.00%;67个带正电的氨基酸残基(Arg+Lys),占总数的12.18%。在大肠杆菌中的半衰期10 h,稳定系数38.40,属稳定的蛋白质类型。AhGPAT5蛋白含的460个氨基酸残基中,Leu(L)和Val(V)含量较高,分别占氨基酸总数的11.7%和11.1%;其次为Arg(R)、Gly(G)和Ser(S),分别占氨基酸总量的7.4%、6.3%和6.3%。含43个带负电的氨基酸残基(Asp+Glu),占总数的9.34%;59个带正电的氨基酸残基(Arg+Lys),占总数的12.83%。在大肠杆菌中的半衰期10 h,稳定系数38.49,属稳定的蛋白质类型。
采用在线HNN数据库对氨基酸序列的二级结构进行预测,结果表明AhGPAT3含α-螺旋185个,占33.64%;β-折叠54个,占9.82%;自由卷曲171个,占31.09%。AhGPAT5含α-螺旋173个,占37.61%;β-折叠50个,占10.87%;自由卷曲139个,占30.22%。
TMHMM在线工具预测发现,AhGPAT3有3个跨膜结构,AhGPAT5有2个跨膜结构,推测AhGPAT3和AhGPAT5均属于跨膜类蛋白。最近有研究认为水稻中的OsGPAT3定位于内质网[12]。保守结构域预测发现AhGPAT3和AhGPAT5基因C端含有一个溶血磷脂酰基转移酶结构域(lysophospholipid acyltransferase,LPLAT)。
在NCBI网站上搜索并下载拟南芥(Arabidopsisthaliana)、大豆(Glycinemax)、花生(ArachishypogaeaL.)GPATs蛋白家族氨基酸序列,对其进行多序列比对分析。结果显示,AhGPAT3氨基酸序列与AtGPAT3、GmGPAT3、AhGPAT2氨基酸序列相似性较高,分别为70%、79%、和78%;与GmGPAT9、AtGPAT9、AhGPAT9氨基酸序列相似性较低,分别为44%、43%和38%。AhGPAT5氨基酸序列与AtGPAT5、GmGPAT5、AhGPAT6、GmGPAT6的氨基酸序列相似性较高,分别为79%、83%、62%、62%;与GmGPAT9、AtGPAT9、AhGPAT9氨基酸序列相似性较低,分别为43%、41%、40%。因此,AhGPAT3和AhGPAT5编码蛋白与其他植物GPAT蛋白序列同源,来自于共同的祖先。
本研究克隆得到的AhGPAT3和AhGPAT5蛋白与其他植物GPAT蛋白的氨基酸序列比对(图3)发现,蛋白序列的相似性很高。两基因都具有典型的酰基转移酶结构域,并且包含四个高度保守的基序AT-Ι、AT-Ⅱ、AT-Ⅲ和AT-Ⅳ。二者都具有催化活性的氨基酸残基组氨酸、天冬氨酸(AT-Ⅰ),甘氨酸(AT-Ⅲ),脯氨酸(AT-Ⅳ);绑定位点氨基酸残基精氨酸(AT-Ⅱ)、谷氨酸(丝氨酸,AT-Ⅲ)[13-14]。

图3 花生与其他植物GPAT蛋白的氨基酸序列比对 Fig.3 Comparison of GPAT amino acid sequences among peanut and other species
注:AT和HAD-like保守结构域分别用方框、直线标出;星号表示HAD结构域中的关键氨基酸残基;三角形表示AT结构域中有催化作用的氨基酸残基;圆点表示AT结构域中的结合位点氨基酸残基。
GenBank登录号:Arachishypogaea(AhGPAT1, JN032676; AhGPAT2, HQ589243; AhGPAT6, HQ589244; AhGPAT8, JX843442; AhGPAT9, JX843441),Arabidopsisthaliana(AtGPAT1, NP_563768; AtGPAT2, NP_563651; AtGPAT3, NP_192104; AtGPAT4, NP_171667; AtGPAT5, NP_187750; AtGPAT6, NP_181346; AtGPAT7, NP_196227; AtGPAT8, NP_191950; AtGPAT9, NP_568925),Glycinemax(GmGPAT1, XP_003545142; GmGPAT2, XP_003520759; GmGPAT3, XP_003536864; GmGPAT5,XP_006575508; GmGPAT6, XP_003529144; GmGPAT8, XP_003520970; GmGPAT9, XP_003533946).
Note: AT and HAD-like domains were boxed (ATI to AT-IV) or marked by lines (HAD-I to HAD-IV), respectively. Critical residues previously identified in similar proteins were marked by asterisks (HAD domain), dots (binding site in AT domain), triangles (catalytic residues in AT domain).
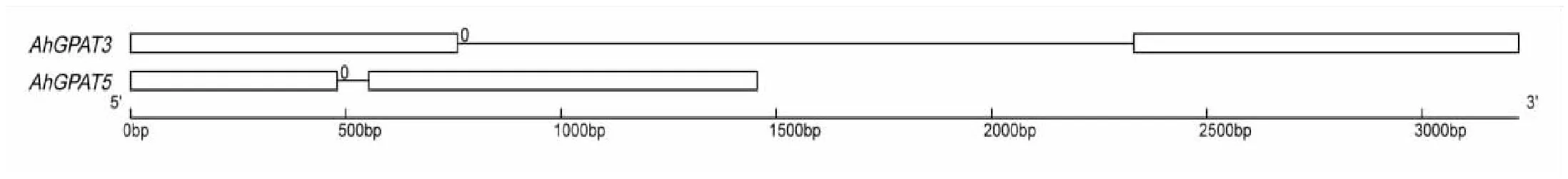
图4 花生AhGPAT3和AhGPAT5的基因结构 Fig.4 The schematic diagram of the structure of AhGPAT3 and AhGPAT5 注:白色方框代表外显子;细线代表内含子;0代表内含子相位。 Note: The white-boxes indicate exon, the line indicate intron, the 0 indicate intron phases.
根据已获得的AhGPAT3和AhGPAT5基因的全长编码序列(CDS),本地BLAST花生全基因组数据库,获得花生AhGPAT3和AhGPAT5的基因组序列。AhGPAT3和AhGPAT5基因组全长分别为3324 bp和1456 bp。利用GSDS工具对两基因进行基因结构和剪接特征分析。结果如图4所示,AhGPAT3和AhGPAT3的基因结构均包括2个外显子和一个内含子。AhGPAT3内含子5'端剪接位点具有AG//GT保守序列,3'端剪接位点具有AG//GA保守序列;AhGPAT5内含子5'端具有AG//G保守序列,3'端具有AG//T保守序列,可知AhGPAT3和AhGPAT5基因外显子和内含子剪接位点的剪接方式符合经典的GT-AG规律[15-16],即内含子5'端剪接位点(受体)具有A(T/C/G)G//GT的保守序列,3'端剪接位点(供体)具有C(T)AG//G(A/T)的保守序列。
2.3 系统发育树分析
如图5所示,对AhGPAT3和AhGPAT5及其他植物GPAT蛋白进行系统发育分析。从树的拓扑结构可以看出,所有的基因聚成了三个不同的分支:ATS1,GPAT1-8,GPAT9。通过对莱茵衣藻(Chlamydomonasreinhardtii)和团藻(Volvoxcarteri)的基因组搜索,发现了叶绿体定位的ATS1和GPAT9基因,但是没有发现其他GPAT家族成员。据报道其他8个GPAT基因可能是植物特有的,在别的物种中不存在[17]。叶绿体定位的可溶性蛋白AhATS1与其他膜结合的GPAT家族成员关系较远。AhGPAT9与高等植物和绿藻的GPAT9蛋白聚在一起,与高等植物的GPAT1-GPAT8蛋白分离开来。GPAT1-GPAT8蛋白分成了3个亚家族。据报道GPAT4/6/8亚家族是最古老的,出现于高等植物进化的初始阶段(苔癣植物)。它们主要参与角质和角质类似物的形成。相反,磷酸酶缺乏的GPAT1-3和GPAT5/7亚家族后期分化产生伴随着维管植物的产生[17]。AhGPAT3基因与高等植物的GPAT2和GPAT3基因聚在一起,AhGPAT5基因与高等植物的GPAT5和GPAT7基因聚在一起。GPAT1-GPAT3亚家族与GPAT4/6/8和GPAT5/7亚家族的亲缘关系较远。因此,AhGPAT3和AhGPAT5基因编码的蛋白虽然都属于GPAT家族,但是二者进化关系却较远,它们在调控植物油脂的合成方面可能具有不同的功能。
2.4 基因的表达特性分析
通过实时荧光定量PCR(qRT-PCR)技术分别检测了AhGPAT3和AhGPAT5基因在花生不同组织器官、种子发育不同时期、3种非生物胁迫条件和6种激素类处理下的表达特性。以花生Actin11(AhACT11)基因作为本实验的内参基因。
图6显示,在花生不同组织中,AhGPAT3基因在下胚轴和花中的表达量较高,分别为子叶和叶中表达量的4.8倍和168倍;在种子发育过程中,发育初期的基因表达量最高,随后表达量明显下降。AhGPAT5基因在花生下胚轴中的表达量最高,约为花中表达量的2.4倍;在种子发育初期基因的表达量最高,发育中后期表达量极低。
图7显示,以0h花生三叶期幼苗为对照,在不同逆境胁迫及激素类处理下,花生叶片中AhGPAT3基因均明显地诱导表达,表明该基因在这些胁迫下对花生抗逆性均有正调控作用。其中,水杨酸处理4h、8h后表达量逐步上调,12h表达量下调后又逐渐上调,72h表达量最高,是对照的25.6倍。过氧化氢处理4h时,表达量上调3倍,8h时表达量下调,之后保持在较低水平。茉莉酸处理24h时表达量最高,为对照的10.5倍。赤霉素处理4h后表达量大幅上调,为对照的4.1倍,随后下降。乙烯利处理24h时表达量显著上调,约为对照表达量的16倍。脱落酸处理后,基因表达量逐渐上调,24h时达最高,是对照的5.4倍,随后显著下降。盐处理4h后表达量最高,是对照的4.5倍,随后又大幅下降,表达量保持在较低水平。低温处理后72 h时,表达量大幅上调,为对照的26倍。干旱处理后12 h时基因表达量最高,为对照的35倍。

图5 植物GPAT基因系统发育树 Fig.5 Phylogenetic tree of plant GPAT isoforms

图6 AhGPAT3和AhGPAT5基因在花生不同组织和种子不同发育时期的表达分析 Fig.6 Expression analysis of AhGPAT3 and AhGPAT5 in six peanut tissues and at five stages of seed development

图7 AhGPAT3基因在不同非生物胁迫条件下的表达分析 Fig.7 Expression analysis of AhGPAT5 under abiotic stresses
图8显示,以0h花生幼苗为对照,低温和过氧化氢处理下,表达量在4 h时大量下调,随后逐渐上调,72 h时表达量最高分别为对照的1.06倍和1.13倍。AhGPAT5基因在高盐、低温和干旱非生物逆境胁迫及激素处理下,表达量均明显下调。表明该基因在非生物胁迫条件下对花生抗逆性可能具有负调控作用。
3 讨 论
提高花生含油量及抗逆性研究对于提高花生产量、缓解我国油料安全具有重要的意义。针对花生油脂合成代谢的关键基因,对花生品种进行定向改良潜力巨大。GPAT蛋白超家族在植物生长发育、角质、木栓质[18]、膜脂和贮存脂质的合成中起至关重要的作用[19],是TAG生物合成途径中的限速酶[20],在植物种子含油量和逆境抗性中有着不可忽视的影响。前人研究表明,拟南芥AtGPAT5基因影响根和种子表皮木栓质的合成[21],在拟南芥和烟草中过量表达AtGPAT5基因,植物叶片表面出现甘油单酯[18,22-23]。Men等[12]研究发现水稻中OsGPAT3功能缺失显著影响花药角质层和花粉外壁的形成,并导致雄性不育。目前,对于花生AhGPAT3和AhGPAT5基因功能的相关研究较少。

图8 AhGPAT5基因不同非生物胁迫条件下的表达分析 Fig.8 Expression analysis of AhGPAT5 under abiotic stresses
本实验克隆得到的AhGPAT3和AhGPAT5编码的蛋白均具有GPAT家族共有的甘油酰基转移酶保守氨基酸序列(AT-I~AT-IV)[24]。因此推测两个基因在功能上可能有与GPAT家族类似的功能,在花生油脂合成过程中起重要作用。AT保守域位于蛋白质序列C端。其中AT-I中的组氨酸和天冬氨酸,AT-III中的甘氨酸,AT-IV中的脯氨酸主要起催化作用。AT-II中的精氨酸和AT-III中的谷氨酸(丝氨酸)主要起绑定3-磷酸甘油底物的作用[25]。除了AT保守域,一个类似卤酸脱卤酶(HAD-I~HAD-IV)的保守域在AhGPAT6和AhGPAT8的N末端被发现。这个保守域一般存在于磷酸水解酶中。HAD-I含有一个典型的DXD框,其中天冬氨酸作为催化过程中的亲核体。HAD-II中的苏氨酸和HAD-III中的赖氨酸有利于反应中间产物的稳定性。HAD-IV中含有酸性氨基酸天冬氨酸残基,有利于Mg2+在活性位点的互作[26]。
实时定量PCR分析发现,AhGPAT3和AhGPAT5基因在下胚轴中表达量最高,说明其可能参与种子萌发。和小燕等[27]研究表明,花生种子发育初期,各类调控脂肪酸合成的基因比较活跃,而随着合成油脂调控的不断继续,相关调节基因表达量下降,同时各类脂肪酸和油脂的合成也渐渐变慢。AhGPAT3和AhGPAT5基因在花生种子发育过程中的表达量变化符合这一现象,初期表达量明显高于其他时期,因此,推测两基因可能参与种子油脂合成。实验下一步计划通过转基因技术将两基因分别转入拟南芥和花生中,阐明两基因在花生油脂合成和抗逆调控中的作用机理。
干旱、低温、高盐等非生物胁迫严重影响植物的生长发育和产量[28-29]。目前,对GPAT基因家族在非生物胁迫方面的研究较少。Szalontai等[30]将拟南芥质体GPAT基因转入烟草中超表达能提高其低温抗性,而将南瓜质体GPAT的cDNA转入烟草中超表达却降低了烟草的抗寒能力。刘继梅等[31]对不同抗冷性水稻品种GPAT保守氨基酸区域进行研究,发现其中3个不同位点的脯氨酸替换与水稻抗冷性相关。Sui等[32]在番茄中超表达LeGPAT基因,提高了其耐低温能力。在本试验为了探究AhGPAT3和AhGPAT5两基因是否对花生各种非生物逆境胁迫和激素处理有响应,并可能参与哪些抗性调节,以期为花生抗性分子调控研究提供新的基因资源。设计了3种非生物逆境胁迫条件及6种激素处理,进行全面调查。结果表明,两基因对多种非生物逆境胁迫有不同的响应模式。AhGPAT3叶片基因受干旱、低温、高盐非生物胁迫明显诱导表达。低温处理72 h后的表达量是对照的26倍,干旱处理12 h后的表达量是对照的35倍,高盐处理4 h后的表达量是对照的4.5倍。因此推测,AhGPAT3基因可能参与了花生非生物胁迫抗性调节过程。胁迫响应基因可能通过两种信号途径调控:ABA依赖型和非ABA依赖型。荧光定量PCR结果表明,在花生叶片中AhGPAT3的表达受ABA诱导明显,推测AhGPAT3以依赖ABA的方式参与非生物胁迫的调控。
本研究从花生中克隆了AhGPAT3和AhGPAT5两个基因。多序列比对和进化树分析表明,AhGPAT3与高等植物的GPAT3和GPAT2亲缘关系最近;AhGPAT5与高等植物的GPAT5和GPAT7亲缘关系最近。荧光定量PCR结果显示,AhGPAT3和AhGPAT5基因在种子发育初期和下胚轴中的表达量较高。另外发现,AhGPAT3基因在花生耐旱、耐低温、耐盐机制中发挥重要作用,很可能参与了花生在干旱、低温和高盐逆境胁迫条件下的抗性调节。
