四倍体杂交冰草SRAP和SSR分子标记遗传连锁图谱构建
2018-08-28杨东升于肖夏李晓宇吴国芳张明飞
杨东升,于 卓,于肖夏,李晓宇,吴国芳,石 悦,张明飞
(内蒙古农业大学农学院,内蒙古呼和浩特 010019)
冰草(Agropyron)为禾本科冰草属多年生草本植物,适应性广、抗旱性和耐寒性强,是牧草及麦类作物杂交改良和遗传转化的宝贵基因资源,具有重要的饲用价值、生态价值和经济利用价值[1]。但由于冰草种间差异的原因,会表现出一定的缺点,如蒙古冰草(Agropyronmongolicum)叶量少、茎叶比高,品质上有待进一步提升[2],而航道冰草(Agropyroncristatumcv. Fariway)则可弥补这些不足。四倍体杂交冰草(2n=4x=28)是本课题组以蒙古冰草为母本,航道冰草为父本,通过种间远缘杂交获得二倍体杂种F1代,用秋水仙碱溶液进行染色体加倍选育而成的优质牧草[3-5],结合了父本航道冰草分蘖性强、叶量大、产量高、营养价值高和母本蒙古冰草抗旱耐寒、耐贫瘠及耐风沙等优点[6]。
SSR(simple sequence repeat)标记具有通用性强、稳定性高、基因组内分布范围广和标记多呈共显性等优点[7-8],常被用于构建植物遗传连锁图谱和分析种质资源的遗传差异性。SRAP(sequence-related amplified polymorphism)标记是由Li等[9]提出的一种新型分子标记,具有操作简便、多态性丰富、价格低廉等优点,可有效增加遗传图谱的分子标记数。SSR和SRAP标记相结合的技术在植物遗传作图研究中已有一些应用,如武耀龙等[10]利用SRAP和SSR标记,成功构建了一张含有128个标记位点的红麻遗传连锁图谱,图谱全长为2155.43 cM;朱洪运等[11]用AFLP、SRAP和SSR标记构建出一张高密度结球甘蓝分子遗传连锁图谱,该图谱含有240个标记,覆盖864.4 cM;郭利建等[12]基于SRAP和SSR标记,通过区间作图法对三原及杨凌两个环境下小麦籽粒的淀粉含量、湿面筋含量、粗蛋白质含量和Zeleny沉降值进行了QTL定位。
本课题组姜志艳等[13]利用AFLP标记首次构建了一张含14个连锁群的四倍体杂交冰草的遗传图谱,覆盖基因组长度2 574.3 cM,标记间平均距离3.82 cM。由于AFLP仅为单分子标记,图谱加密有限,且预扩增时需进行酶切和连接等过程,其操作复杂且成本高;而SSR与SRAP标记相结合,可使遗传图谱上的标记分布更为均匀、精度提高[14]。目前关于SRAP和SSR标记在四倍体杂交冰草遗传作图上的研究尚未见报道。本研究以四倍体杂交冰草F2代180个分离单株及其亲本为作图材料[13,15],利用SRAP和SSR标记技术构建其遗传连锁图谱,以期为冰草AFLP、SRAP和SSR三种标记整合的高密度遗传图谱构建,以及产量品质、抗旱耐寒等重要性状的QTL精细定位、功能基因图位克隆和分子标记辅助育种研究奠定基础。
1 材料与方法
1.1 供试材料
供试材料为四倍体杂交冰草180个F2单株及母本蒙古冰草和父本航道冰草。各材料均种植于呼和浩特市赛罕区内蒙古农业大学农作场。
1.2 DNA的提取与纯度检测
在分蘖期分别取杂交冰草F2代180个单株及亲本的幼嫩叶片,剪碎混匀,各取0.15 g于研钵中加液氮充分研磨,利用植物基因组试剂盒(天根生化科技(北京)有限公司)提取DNA,利用1.5%的琼脂糖凝胶电泳检测DNA纯度。将符合要求的基因组DNA稀释至80 ng·μL-1,置于-40 ℃ 冰箱保存备用。
1.3 SSR和SRAP的引物来源
本试验所用的108对SSR引物来自NCBI (https://www.ncbi.nlm.nih.gov/probe/)公布的小麦(Tritiumaestivum)、高粱(Sorghumbicolor)及玉米(Zeamays)上已开发的SSR引物。SRAP引物来源于Li等[9]、Pezzotti等[16]及Lin等[17]已报道的引物,其中正向和反向引物均为12条,将其自由组合得到144对引物,委托生工生物工程(上海)股份有限公司合成。
1.4 反应体系及程序
SRAP-PCR体系参考Taq polymerase(纽英伦生物技术(北京)有限公司)说明书,扩增程序为94 ℃预变性5 min;94 ℃变性55 s,35 ℃复性1 min,72 ℃延伸1 min,5个循环;94 ℃变性1 min,50 ℃复性1 min,35个循环;72 ℃延伸8 min,4 ℃保存备用。SSR-PCR体系与程序参考Taq polymerase(纽英伦生物技术(北京)有限公司)说明书,其中循环数为35,退火温度为57 ℃,延伸时间为1 min。参照石 悦等[18]的方法对扩增产物进行电泳检测。
1.5 数据统计
多态性位点统计:对SRAP及SSR多态性标记位点进行记录,有条带的记录为“1”,无条带的记录为“0”,缺失或条带不清楚的记录为“-”,生成“0”和“1”组合的原始矩阵。多态性位点百分率:P=(K/N)×100%,其中K表示多态性位点数目、N表示测定位点总数。
偏分离统计:根据孟德尔分离规律,四倍体杂交冰草F2群体的基因型理论上符合3∶1的分离比,在0.05和0.01的水平上,利用卡方检验SRAP和SSR标记多态性位点,判断每一个被检测到的多态性位点是否发生偏分离。
1.6 遗传连锁图谱构建
将统计的SRAP和SSR标记位点数据中的“1”和“0”矩阵转换成软件要求的目标格式,利用作图软件Join Map 3.0构建四倍体杂交冰草F2分离群体的遗传连锁图谱。
2 结果与分析
2.1 SRAP和SSR引物筛选及分子标记多态性分析
以亲本及F2群体中的任意3个子代DNA为模板进行适宜引物筛选,在144对SRAP引物自由组合中,共筛选出了10对条带清晰、多态性高的引物(图1,图2)。这10对SRAP引物组合共扩增出267个位点,其中多态性位点为240个,平均每对引物产生24个多态性位点,多态性位点百分率为89.9 %(表1)。
用108对SSR引物对亲本及F2子代进行检测,共得到10对SSR多态性明显且带型清晰的引物组合(图3,图4)。这些引物组合共产生了277个位点,其中多态性标记位点为235个,平均每对引物组合扩增出23.5个多态性标记位点,多态性位点百分率为84.8 %(表2)。

P1:航道冰草;P2:蒙古冰草;1~3:F2单株。P1:Agropyron cristatum cv. Fariway; P2:Agropyron mongolicum; 1-3:F2 plants.
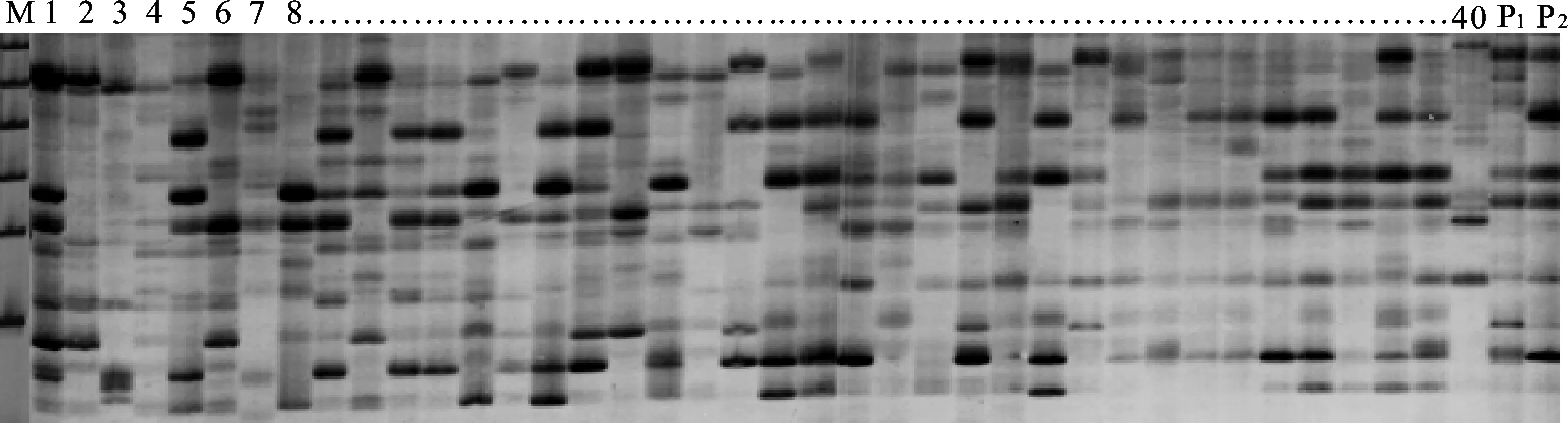
P1:航道冰草;P2:蒙古冰草;1~40:F2单株群体部分单株P1: Agropyron cristatum cv. Fariway; P2:Agropyron mongolicum; 1-40:Partial F2 individuals.

P1:航道冰草;P2:蒙古冰草;1~3:F2单株。P1:Agropyron cristatum cv. Fariway; P2:Agropyron mongolicum; 1-3: F2 plants.

图3 四倍体杂交冰草部分SSR适宜引物的筛选结果Fig.3 Screening of suitable SSR primers for tetraploid hybrid crested wheatgrass
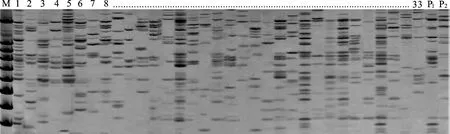
P1:航道冰草;P2:蒙古冰草;1~33:F2单株群体部分单株。P1:Agropyron cristatum cv. Fariway; P2:Agropyron mongolicum; 1-33:Partial F2 individuals.
2.2 标记偏分离分析
对获得的共475个标记位点进行卡方检测结果显示,在0.05水平上,其中不符合孟德尔分离比的位点有86个,占总标记数的18.11%。四倍体杂交冰草的14个连锁群均分布有偏分离标记,偏分离标记数的范围为2~11,偏分离标记比率的范围为13.25%~23.81 %,符合植物遗传作图的偏分离比率小于30%的要求(表3)。
2.3 遗传连锁图谱的构建
利用软件Join Map 3.0进行遗传连锁图谱的构建。在LOD≥3.0的条件下,构建出一张包含475个标记(240个SRAP标记+235个SSR标记)、14个连锁群的四倍体杂交冰草遗传连锁图谱,覆盖基因组总长度为1592.7 cM,标记间的平均图距为3.35 cM,各连锁群长度范围为67.6~145.2 cM,标记数目在10~83之间,其中标记数目最多且最长的连锁群为LG2,标记数目最少且最短的连锁群为LG8(表4,图5)。

表3 四倍体杂交冰草作图标记的偏分离分析Table 3 Segregation distortion analysis of mapping markers of tetraploid hybrid crested wheatgrass

表4 四倍体杂交冰草遗传图谱的基本特征Table 4 Basic characteristic of the genetic linkage map of tetraploid hybrid crested wheatgrass

*表示偏分离标记。 * represents distorted marker.
3 讨 论
高质量的遗传连锁图谱是QTL精细定位、基因图位克隆及分子标记辅助育种的有效工具[19-20],因此提高连锁图谱的质量显得尤为重要。图谱的质量主要取决于分子标记位点在连锁群上分布的均匀程度,而均匀程度又受限于标记类型,将不同类型的分子标记相结合构建图谱,会显著提高标记在图谱上分布的均匀程度,并有利于克服不同分子标记间的缺陷[21]。例如,金梦阳等[22]通过SRAP、SSR、AFLP和TRAP四种标记构建了一张包含20个连锁群,共300个标记位点的甘蓝型油菜遗传图谱,且各标记在图谱的不同连锁群上分布较均匀。Hashizume等[23]利用477个RAPD标记、53个RFLP标记及23个ISSR标记,构建了西瓜的遗传连锁图谱,标记位点分布均匀。以上这些标记为第一、二代分子标记,其最大优点是操作简便、且电泳条带真实可见,可直接用于作物相关性状QTL的精细定位研究;其不足点是提供的遗传信息量有限。随着近年来基因组测序技术的改进,SNP分子标记技术倍受关注,该标记技术的主要优点是信息检出量大,易于基因分型;其不足点是SNP等位基因位点在多倍体植物的基因组中的比例具有可变性,很难完成单一型的鉴定,且成本高。
本试验所用材料为四倍体杂交冰草,基于SRAP标记主要对编码区进行序列扩增,而SSR标记的大部分扩增区域位于非表达区[24]的特点,同时使用这两种标记技术对其进行遗传作图,可减小标记位点在图谱上分布的不均匀程度。研究表明,SRAP和SSR两种标记位点在四倍体杂交冰草的每个连锁群上分布较均匀,图谱整体质量有所提高,这为下一步整合AFLP、SRAP和SSR三种标记构建四倍体杂交冰草的高密度遗传图谱奠定了基础。
偏分离现象被认为是生物进化的推动力,在植物界中普遍存在[25]。目前已在大麦[26]、水稻[27]和番茄[28]等作物中发现了偏分离现象 。偏分离比例会因物种和作图群体类型的不同而有所变化,有多种因素可造成遗传标记产生不同程度的偏分离现象。Perfectti等[29]认为偏分离是由配子体和合子体共同选择作用引起的;Pawlowshi等[30]的研究表明,环境因素、基因转换及转基因沉默均可能造成偏分离现象的发生。偏分离标记会在一定程度上通过影响标记间的距离来影响图谱的质量,植物遗传作图时通常要求偏分离标记比例小于30%[10]。本试验得到的475个标记中,有389个标记符合孟德尔分离规律,有86个标记发生偏分离,这些标记不规则地分布于14个连锁群上,各连锁群的偏分离标记比例变幅在13.25~23.81 %之间,平均偏分离比例为18.11 %,可进行四倍体杂交冰草的遗传作图。
