云南元江上游石羊江河谷绿孔雀夜宿地选择
2018-08-21刘钊杨爱芳谢宗平谢以昌谢全刚王进张仁功黄庆文文云燕周伟蒋志刚
刘钊杨爱芳谢宗平谢以昌谢全刚王进张仁功黄庆文文云燕周伟蒋志刚
(1.河西学院农业与生物技术学院,张掖,734000;2.中国科学院动物研究所动物生态与保护生物学重点实验室,北京,100101;3.西南林业大学保护生物学学院,昆明,650224;4.云南省双柏恐龙河自然保护区,双柏,675107;5.云南省楚雄州自然保护区管理局,楚雄,675000)
栖息地选择是指动物对其存活所必需的生境特征和资源的选择过程[1],动物对某种资源往往有较高的选择概率[2-3]。对栖息地的偏好和选择有助于提高动物的适合度[4]。研究栖息地选择可以为动物生境保护和管理提供理论依据,因此,人们侧重研究探讨动物的生境特征和生境质量,并确定生境特征与动物适合度之间的关系。
绿孔雀(Pavomuticus)在全世界记录有3个亚种,其中P.m.imperator分布于缅甸东部、中国云南、泰国和中南半岛[5]。绿孔雀野生种群数量稀少,已被列为国家Ⅰ级重点保护动物。由于人类经济活动的影响,其栖息生境被破坏,种群数量明显下降,分布区域急剧缩小,导致现存种群形成小家族群点状隔离分布[6]。对越南南部绿孔雀生存现状和栖息地利用研究表明,绿孔雀偏好选择接近永久性水源地和无人为干扰的生境,居民点的扩增、栖息地丧失和水源的可获得性是其主要限制因素[7]。
夜宿地是动物用来夜间休息的场所,其质量的好坏关系到动物夜间的安全。夜宿地是鸡形目(Galliformes)鸟类重要栖息地之一。已有研究发现鸡形目鸟类对夜宿地有明显的选择性[8-9]。本文期望通过对云南元江上游石羊江河谷绿孔雀极小种群夜宿地选择的研究,确定影响夜宿地选择的关键性生态因子,揭示环境特征对绿孔雀栖息地选择的影响,解释其濒危成因。同时分析生境破碎化和人为干扰对绿孔雀生存的影响,为绿孔雀极小种群保护方案的制定提供依据。
1 研究地点与方法
1.1 研究地概况
研究区位于元江(红河)上游石羊江河谷,处于云南楚雄州双柏恐龙河自然保护区内(N 24°56′~26°09′,E 98°34′~98°50′),系哀牢山系云岭支脉,地处元江大断裂带。因江河沿断裂带下切,谷低箐深,地势险峻,形成河谷和低山两种地貌类型。保护区最高海拔1796 m,最低海拔623 m,相对高差1173 m。该区气候属西部高原季风气候,5~10月为雨季,11月至翌年4月为旱季。石羊江河谷处于季风的背风面,地形的焚风效应造成河谷底部的干热生境,年均温度14.0 ℃,年均降水量924.6 mm,年均日照2355 h[10]。具体研究区位于保护区南部的磨家湾,最低海拔650 m,最高海拔1370 m,面积约16 km2。小江河流经研究区南部,并与石羊江在该区东部交汇。除有绿孔雀以外,研究区内雉科鸟类还有黑颈长尾雉(Syrmaticushumiae)、原鸡(Gallusgallus)和白腹锦鸡(Chrysolophusamherstiae)等。
1.2 栖息地调查
野外工作时间为2007年春季3月15日~4月15日和秋季10月25日~11月25日。通过访问保护区工作人员和当地居民,结合实地查证,以了解绿孔雀的活动范围、时间、地点和痕迹特点(包括粪便、羽毛和啄痕)。观察发现,绿孔雀在6:00~8:30和17:00~19:30两个时段鸣叫频繁,阴天或雨后更加频繁。在绿孔雀频繁活动区,经常能发现粪便和羽毛。绿孔雀的粪便有2种,一种具白色尿酸结晶,螺旋状,较其他雉类大,长度为3~5 cm;另一种为黑色,锥状或块状,不呈螺旋状,较大,长度为4~7 cm,新鲜的黑色粪便常伴有恶臭味。收集到的羽毛多浓褐色泛绿辉,为孔雀背羽。雉类常啄食落果,留下明显的啄痕,绿孔雀的啄痕较其他雉类大。
采用样线法调查种群数量和空间分布,为保证样线均匀分布在整个研究区域和穿越绿孔雀栖息的不同生境,共设置6条不重叠的样线,样线单侧宽30 m,基本为调查人员的可视距离,每条样线长4~5 km,以计步器测定。于绿孔雀活动高峰期(6:00~9:30;17:00~19:00)内,以2~3 km/h 的速度行走样线,每隔3~4 d重复行走样线,每条样线重复5次以上。观察绿孔雀的实体及活动行为,搜索活动痕迹(包括新鲜粪便、羽毛和啄痕),并作短距离的循迹搜寻,记录所见实体的数量和所在活动位点的经纬坐标。以直接观察到的绿孔雀的采食行为、啄痕并结合观察到的粪便痕迹确定取食位点。夜宿树的确定方法:(1)直接观察法:以直接观察到的绿孔雀的夜宿树确定。(2)守候寻找法:据观察结果,确定大致夜宿时间(19:00~19:30),在可能夜宿的地域守候。绿孔雀上树夜宿前常发出鸣叫,根据鸣声判断夜宿树的大致方位。第二天清晨,通过寻找树下的新鲜粪便确定夜宿树。(3)访问法:访问当地放牧居民,由他们提供近期绿孔雀的夜宿地点。
根据野外观察结果,以夜宿树为中心,设置10 m×10 m的大样方为夜宿地样方,在每个大样方内对角线的4等分处设置1 m×1 m的小样方4个[11]。以大样方作为乔灌木因子的生境测度,小样方作为草本、落果和种子等因子的测度,共调查并确定了19个生态因子[11](表1)。同时,以样方中心为参照点,通过随机数字表确定对照样方的中心,设置相同大小的对照样方[12],对照样方经判断均未被绿孔雀利用作为其夜宿地点。随机数字为5位数,前 3位为利用和对照样地中心点之间的方位角,后2位作为二者中心点的距离。当前3位的数值超过360时,减去360的整倍数后的余值作为方位角。以正北为0°起点,顺时针旋转罗盘,使指针指到方位角值,此时的方向即为对照样方的位置方向[13]。对照样方测量的因子和方法同利用样方。
表1 绿孔雀夜宿地利用与对照样方之间的生态因子比较

Tab.1 Comparisons of means of green peacock roosting habitat variables of different sampling plots
AL:altitude;SL:slope;POS:position on slope;DT:density of trees;CT:coverage of trees;TT:type of trees;AHT:average height of trees;ADCT:average diameter at the chest-height of trees;DS:density of shrub;CS:coverage of shrub;AHS:average height of shrub;CG:coverage of grasses;DF:density of fruits;TS:thickness of shatter;DSW:distance to water;DR:distance to road;DSC:distance to countryside;DFE:distance to forest edge;DL:density of Liana
在“t(z)”栏黑体斜排的数据为非参数检验结果 The bold and italic data in columnst(z)are results of nonparametric tests.*P<0.05;**P<0.01
1.3 数据处理
1.3.1 生态因子比较和判别分析
比较夜宿地利用样方与对照样方间坡向因子的差异显著性,先检验数据的均匀性,若存在集中趋势,采用独立样本t检验;否则,采用非参数的U检验。因坡位因子为定性因子,分为上、中和下坡位,为量化分析依次赋值3、2、1。采用单样本的 Kolmogorov SmirnovZ检验分析其他生态因子的正态性,当数据符合正态分布时,采用配对样本的t检验;否则,采用非参数的 Wilcoxon 符号秩检验,以比较生态因子的差异[12]。采用判别分析,比较秋季利用样方与对照样方间主成分分值的差异,以分析不同样地间的分化[12]。
1.3.2 逻辑斯蒂回归分析

AICc=AIC+2K(K+1)/(n-K-1)
式中,K为回归等式中自变量的个数加2,n为样本总数。
对所得模型进行 Hosmer & Leweshow检验[15],确定模型对因变量变化的判别是否达到显著水平(P>0.05)。在模型的切断点(cut-off point)为0.5时,用计算模型的预测值与实际观测值的匹配系数的方法来验证模型的准确性,统计并计算利用样方和对照样方的正确预测数和正确预测率[17]。
2 结果与分析
在研究区内,春、秋季共设置夜宿地利用样地与对照样地各46个。最低样方海拔为 750 m,最高样方海拔为 1155 m。
2.1 生态因子比较和生态位分化的判别


图1 绿孔雀夜宿地利用样方和对照样方典则判别函数值分异图Fig.1 Histogram for canonical discriminant functions’ valuewhich resulted from different sampling plots inthe roosting habitat of green peacock
2.2 逻辑斯蒂回归分析与概率预测
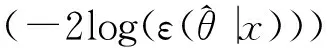
表2 绿孔雀对夜宿地选择的逻辑斯蒂回归分析结果

Tab.2 The results by logistic regression for roosting sites of green peacock Pavo muticus
注:模型切断点为0.5;表中英文缩写同表1
3 讨论
3.1 绿孔雀对夜宿地的偏好
影响绿孔雀夜宿地选择的重要因素为地形和隐蔽条件。因子的差异性检验和逻辑斯蒂回归分析表明,绿孔雀主要选择海拔较高、坡度较大、隐蔽条件较好的山脊夜宿。鸟类能识别环境中的某些特征,并根据这些特征主动选择生活环境[18]。研究发现,绿孔雀的夜宿地多为乔木胸径和盖度较大的林区,乔木的隐蔽条件是躲避敌害的天然屏障,降低了捕食风险。观察发现,绿孔雀夜宿树胸径和盖度较大,均长有横向的粗壮分支,能提供较好的隐蔽和栖位。海拔较高的山脊周边较为空旷,有利于绿孔雀向山谷飞行以躲避敌害。观察亦发现,绿孔雀有沿着海拔梯度向上行走觅食的习性,至傍晚多到达海拔较高处,随即找适宜的安身处夜宿。而较丰富的落果和藤本密度将有助于绿孔雀在夜宿前补充食物和登高夜宿。可见,绿孔雀对夜宿地的选择最终可以体现在某些关键性生态因子上,这些关键性生态因子影响着绿孔雀对栖息地的选择。鉴于到本研究未调查夜宿树的树种信息,期望在今后的绿孔雀栖息地植被群落专项研究中能进一步查明。
3.2 栖息地的分化与行为策略
绿孔雀的夜宿地与对照样地相比,存在着明显的分化。判别分析表明,差异性较大的因子直接决定了栖息地的分化,这与绿孔雀对栖息地的选择偏好及其生存策略有关。而前期的研究结果表明,绿孔雀的觅食地一般海拔较低、食物资源丰富且隐蔽条件好[19-21],相比之下,夜宿地的环境因子与之不同。动物栖息地选择行为表明可供选择的栖息地之间存在差异,而这些有差异的栖息地恰好为野生动物提供了不同的生态环境,从而影响它们的生存和繁衍[18-20,22]。绿孔雀的夜宿地多位于山脊地区,两侧海拔较低。研究结果亦表明,夜宿地的海拔高度显著低于觅食地[19],相比之下,夜宿地的环境特征与对照样方存在明显差异(图1)。前期的调查发现,绿孔雀在觅食时如果在下方遇到人为干扰,逃逸时常向上奔跑,然后迂回朝向海拔较低、草灌木密生的山谷地区,这有利于加速逃跑和寻找隐蔽[19];绿孔雀有日间在低海拔觅食,傍晚上行到海拔较高处夜宿的习性,夜宿地较好的隐蔽条件并位于山脊地区将有利于其躲避敌害时向山谷飞行,或飞至位于山谷的觅食地觅食。调查还发现,夜宿地常有粪便累积,表明在无人为干扰的情况下,绿孔雀常在同一区域内选择夜宿点。绿孔雀选择海拔较高处夜栖,是在获取食物和躲避敌害之间的一种权衡。栖息地的分化,能为生存策略的实现提供了前提条件,绿孔雀的分布及栖息地选择策略与栖息地环境特征和空间资源的异质性程度有关[20-22]。
绿孔雀主要分布在磨家湾的河谷地区,有居民点和林区公路分布在河谷之上的海拔较高处,人为干扰对绿孔雀的影响较大。村民饲养的家畜一般会被定期驱赶至山下河谷觅食,绿孔雀的利用生境几乎都有放牧羊群踩踏、啃食和排便的痕迹。同时河谷上方北侧的山体陡峭,滑坡严重且伴有泥石流发生,这对绿孔雀的生境造成了直接的破坏。又由于该地区蕴藏矿产,探矿和频繁的车辆运输均对绿孔雀的生存造成压力。鉴于保护区内的绿孔雀种群为濒危极小种群,建议相关部门应当及早采取有效的措施,严禁采矿行为,减少人为放牧干扰,恢复脆弱的生态环境,以促进绿孔雀种群的复壮。
致谢:野外调查得到云南楚雄州恐龙河自然保护区全体工作人员的大力协助,磨家湾赵同荣、赵家昌一家提供了食宿条件,在此一并表示感谢!
