依据白桦双列杂交子代林对优良碳汇亲本及家系选择1)
2018-08-13纳晓莹王秀伟刘桂丰姜静
纳晓莹 王秀伟 刘桂丰 姜静
(东北林业大学,哈尔滨,150040)
森林是陆地生态系统中最大的碳库之一,在维护全球气候稳定,调节全球碳循环以及减缓CO2等温室气体浓度上升等方面具有不可替代的作用[1]。森林碳库约占全球植被碳库的86%以上[2-3]。当增加森林密度和区域时,森林就成为了CO2的“汇”。基于联合国环境规划署和粮农组织估计,亚洲森林每年每公顷大约吸收5 000 kg CO2。我国森林的碳储量主要集中于云杉林、冷杉林、落叶松、栎类林、桦木林、硬叶阔叶林和阔叶混交林等7个林分类型中,其碳储量约占整个森林的70%,且中、幼龄林较成熟林,其面积和碳储量都呈上升趋势,如果对现有森林加以更好的抚育管理,作为CO2的“汇”,我国森林还有很大的潜力发挥其碳汇功能[4]。东北地区森林资源面积为6×1011hm2,占全国森林总面积的28.1%,是我国最大的林区[5]。但东北森林的遗传改良较晚,且蓄积量较小,碳储量较低,碳汇的提升空间很大,应寻求更多更有效的方法提高东北森林的碳汇能力,增加其碳储量。
白桦是广布亚欧大陆的世界性树种,在我国东北地区蓄积量较大。近些年来,随着育种技术的发展,研究者对白桦进行多层次的评价选择[6-7],自20世纪90年代起进行了多次种源试验,得到了高碳汇优良种源草河口、清源、帽儿山和小北湖,其地上部分碳储量较对照提高了17.3%~47.2%[8],并对白桦进行优树选择、扦插[9]、强化育种技术[10]的研究,并利用杂交育种手段[11-13]、遗传转化等方法培育出许多速生优质的白桦新种质。其中,杂交育种是培育白桦优良品种的重要手段,杂交后代的基因重组可产生杂种优势[14]。双列交配设计作为杂交设计的主要方法之一[15],目前已广泛被应用于林木遗传改良中[16-21],配合力则是决定杂种具有亲本优势强弱的重要因素,是选配亲本的主要指标。目前,林木遗传改良工作缺乏以选育碳汇良种为目标的研究,因此,应将白桦种质资源改良与碳储量联系起来,筛选固碳能力强且生长性状优良、抗逆性强的白桦种质资源,这样既可以提高白桦人工林生产效率,又可以为全球环境的改善做贡献。
本研究以4株中国白桦优树和1株欧洲白桦优树为杂交亲本,按双列交配设计所得的全同胞子代为研究对象,对其生长性状和碳含量进行测定,分析其碳储量及亲本的一般配合力、特殊配合力和反交效应,旨在对高碳汇的白桦家系及优良杂交组合进行评价。
1 试验地概况
试验地点在东北林业大学帽儿山实验林场林木改良实验站北林13林班(E127°30′~127°34′,N45°20′~45°25′),海拔358 m。土壤为暗棕壤,西南坡向,坡度为15°。该区域为大陆性季风气候,有典型的四季交替,冬季长,夏季短。冬季寒冷干燥,夏季炎热湿润,少风多雨。年平均气温2.8 ℃,平均年降水量723.8 mm。
2 材料与方法
实验材料为7年生白桦双列杂交子代测定林,亲本为4株白桦优树(5、8、11、15号)和1株欧洲白桦(14号)。2005年4月下旬按照双列杂交第3种方式进行控制授粉,获得全部20个正交和反交组合的杂交种子(白桦自交不育),于2006年育苗,2007年5月定植,试验设计采用随机完全区组设计,20个杂交组合,3个区组,区组与区组之间用落叶松间隔。每小区18株,双行排列,株行距2.0 m×2.0 m。2012年进行全林调查,每木检尺,根据调查结果在每个小区中选择1株平均木进行生物量测定,每个家系3株作为样木。
2.1 生物量的测定
本研究中生物量的测定是通过对20个白桦全同胞家系分别将样木伐倒,测量树皮、去皮树干(以下树干均为去皮树干)、枝、叶、根的鲜质量和干质量。树干和树皮的鲜重是将样木按照1m区分段,剥皮并测定各区分段的去皮树干和树皮的鲜质量;枝叶的鲜重是通过选取标准枝,分别称枝和叶的鲜质量;根部的测定采用全挖法,分别测定大根(直径d>5 mm)、粗根(直径在2 mm≤d≤5 mm)、细根(直径d≤2 mm)的鲜质量。干质量的测量是对每株样木的地上部分各区分段的树皮、去皮树干和枝、叶、根分别取样,将所采集的样品带回室内烘干并测定其质量,通过其含水量,计算出样木各组织部位的干重,即各组织部位的生物量,求和得到单株生物量。地上部分生物量指树皮、树干、枝、叶的生物量总和,地下部分生物量指3部分根的生物量总和。
2.2 碳质量分数的测定
碳质量分数的测定采用Multi N/C3000分析仪连用HT1500固体模块(Analytic Jena AG.,Germany)。具体方法:用分析天平分别称取20 mg各组织部位的烘干粉末样品,置于HT1500的燃烧炉中进行燃烧,燃烧温度是1 200 ℃,每个样品的测量时间控制在2~3 min。燃烧过程中有高纯氧气不断通过,确保样品在99%的高纯氧气中充分燃烧。电脑分析计算产生CO2的量来确定样品的全碳含量。
2.3 数据处理
本研究中,各组织部位碳储量和单株碳储量所依据的计算方法如下:
式中:Ct为单株碳储量;Ci为第i个组织部位的碳储量;Bi为第i个组织部位的生物量;Pi为第i个组织部位的碳质量分数。
采用Excel2010(Microsoft)对数据进行整理,使用SPSS19.0统计分析系统对家系间进行方差分析,若方差分析差异有统计学意义,则用Duncan法进行组间多重比较。
采用ForStat2.1统计分析系统进行遗传效应分析[22],根据固定模型进行方差分析,在差异显著的基础上估算双列杂交设计的一般配合力(GCA)、特殊配合力(SCA)和反交效应值(REC)[19],并对配合力进行多重比较和优良亲本的选择。根据随机模型估计一般配合力方差、特殊配合力方差(表1)[23],并用下列公式计算遗传力。

表1 配合力和反交效应值方差分析模型
注:p是亲本数,f是杂交组合数(家系数),f=p(p-1),b是区组数,Vg是一般配合力方差,Vs是特殊配合力方差,Vr是反交效应方差,Ve是误差方差。
加性方差VA=4Vg,显性方差VD=4Vs,遗传方差VG=VA+VD,表型方差VP=VA+VG+Ve,狭义遗传力h2=VA/VP,广义遗传力H2=VG/VP。
3 结果与分析
3.1 生物量和碳质量分数的差异显著性
对白桦双列杂交子代单株生物量及各器官进行方差分析结果表明(表2):在不同杂交组合产生的子代家系间单株生物量达极显著差异水平(P≤0.01);地上部分生物量达极显著差异水平,而地下部分差异不显著;地上部分各组织部位生物量除树皮外,其它差异均达极显著水平;各组织部位的碳质量分数中,除树皮和树枝无显著差异外,其它均有极显著或显著的差异。由于各组织部位样品的碳质量分数并不能代表此部分所存储的碳总量,因此要乘以此组织部位的生物量得到其碳储量估计值,作为此部位的碳储量值。因为碳储量与碳质量分数以及生物量密切相关,后两者的差异显著性会影响碳储量的差异显著性。
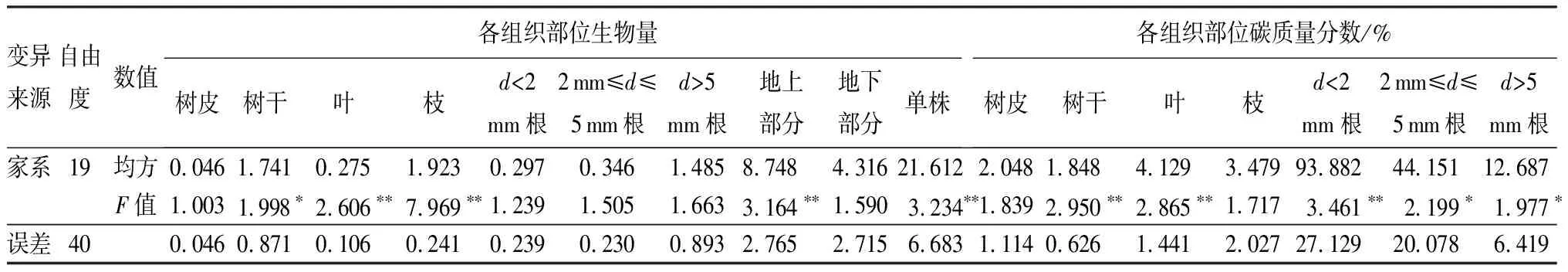
表2 白桦全同胞子代生物量、碳质量分数方差分析
注:** 表示在0.01水平上差异极显著;*表示在0.05水平上差异显著。
3.2 树高、胸径和碳储量的差异显著性分析
白桦杂交子代各树高、胸径、碳储量的变幅及变异系数分析(表3)显示:白桦杂交子代的树高、胸径、单株碳储量的变异系数分别为11.09%、15.39%、28.71%,各组织部位碳储量的变异系数较高,各组合产生的家系性状间存在丰富的变异,表明根据生长性状进行优良杂交组合的选择潜力很大,并且选择高碳汇能力白桦家系的可能性很大;对白桦双列杂交子代树高、胸径、单株碳储量以及各组织部位碳储量进行方差分析(表4)表明:在不同杂交组合产生的家系间除树高、皮碳储量和根碳储量差异不显著,胸径及其他各部分、单株碳储量差异均达到极显著水平(P≤0.01),为筛选优良的碳汇家系提供可能。
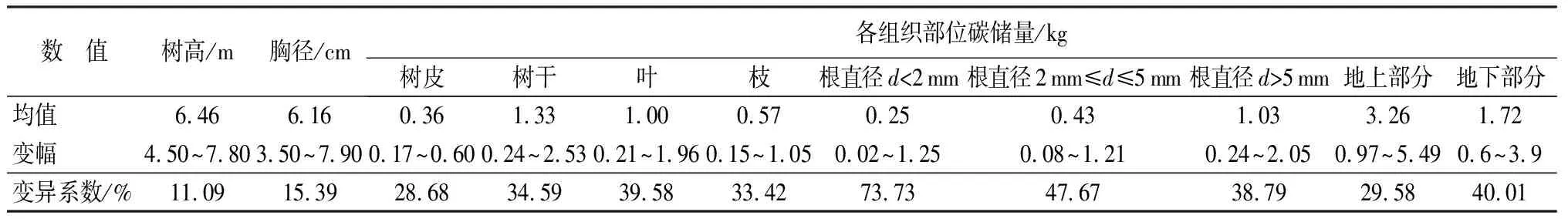
表3 树高、胸径及碳储量变异分析
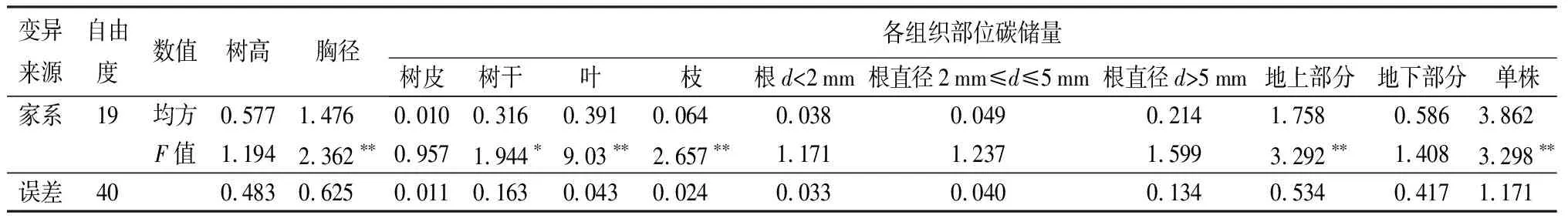
表4 白桦全同胞子代树高、胸径及碳储量方差分析
注:** 表示在0.01水平上差异极显著;*表示在0.05水平上差异显著。
对不同家系单株碳储量进行多重比较(表5),家系619和643最高,平均值分别为7.14和6.89,分别比总体平均值高43.67%,38.63%。627、637、602、642、643和619等6个家系平均值较高,单株碳储量较各家系平均值高13.69%~43.67%,将产生这些家系的杂交组合11×15、14×11、5×8、15×8、15×11、8×14定为待选高碳汇优良亲本组合。以上6个杂交组合中,变异系数较大的组合为14×11和15×11,其单株碳储量变异系数分别为21.54%和23.80%;其次为11×15和5×8,变异系数分别为19.65%和17.11%;变异系数较小的组合为8×14和15×8,分别为7.14%和8.88%。以上数据表明不同的杂交组合,其杂种子代的变异程度是不同的,可根据变异程度对各杂交组合进行遗传评价,全同胞家系单株碳含量变异越明显,就越有利于根据需要进行遗传选择。
3.3 一般配合力、特殊配合力、反交效应分析
配合力是用于阐明子代性状因亲本配合所表现的遗传能力,它表达亲本组合对子代表现的量化遗传关系,是遗传评价的重要参数[15]。根据固定模型进行方差分析,在差异显著的基础上估算双列杂交设计的一般配合力(GCA)、特殊配合力(SCA)和反交效应值(REC)。特殊配合力效应值为正,表明该组合的表现优于双亲的平均表现,反之则劣于双亲的平均表现。因此,分析亲本的一般配合力和特殊配合力,是筛选优良杂交组合的依据之一。反交效应是选择优良组合的正反交组合的要参考依据。若反交效应为正向效应,表明其亲本在正交情况下一般配合力提高了后代在相应性状上的表现,则要选择正交组合;若反交效应为负向效应,表明其亲本在正交情况下一般配合力降低了后代在相应性状上的表现,则选择反交组合。
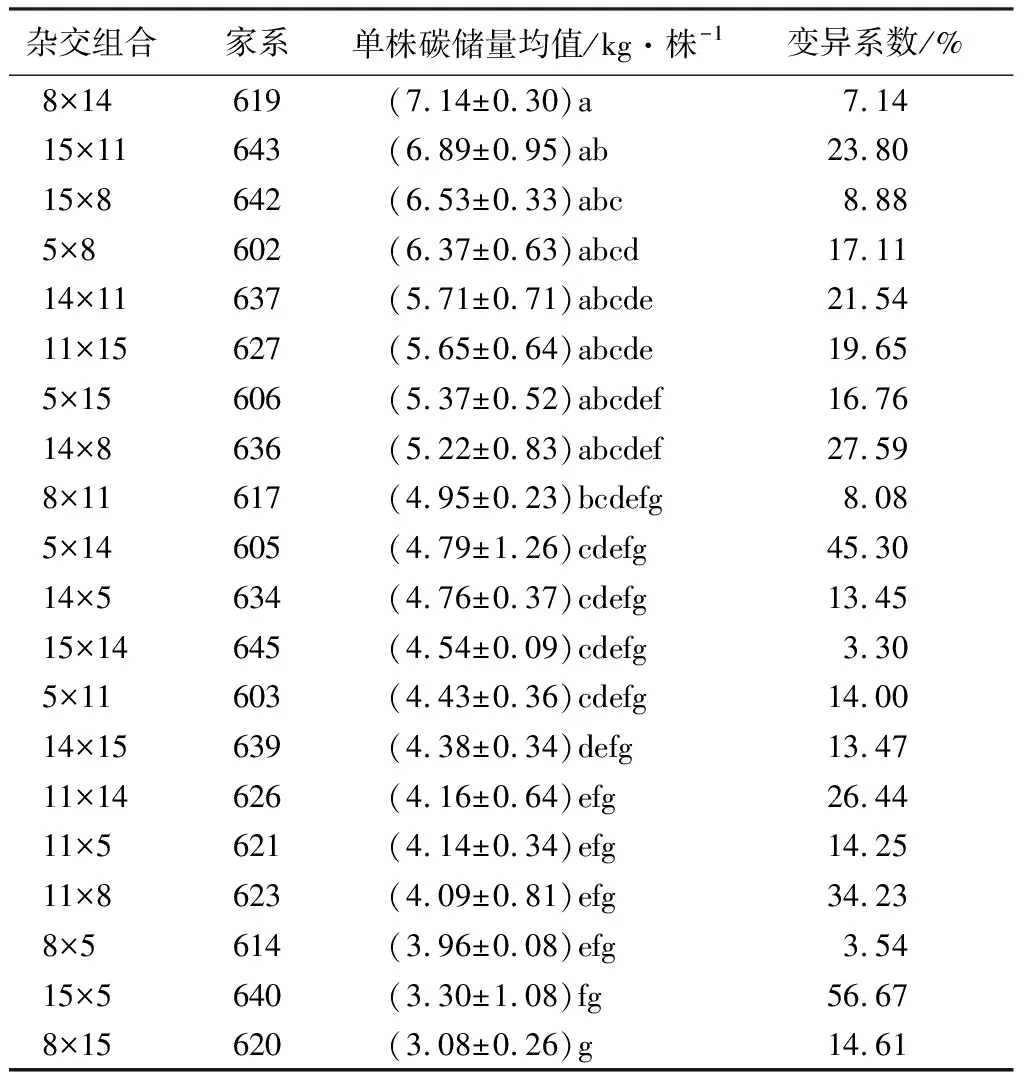
表5 白桦全同胞子代单株碳储量多重比较
注:表中数据表示平均值±标准差;同列不同字母表示在0.05水平上差异显著。
根据固定模型对白桦各杂交组合的碳储量进行配合力和反交效应方差分析,结果显示:单株碳储量的一般配合力差异不显著,特殊配合力和反交效应均达到了的极显著水平(表6),遗传力达到70%以上,表明特殊配合力和反交效应值存在显著的遗传差异,可根据它们进行下一步的估算和分析。
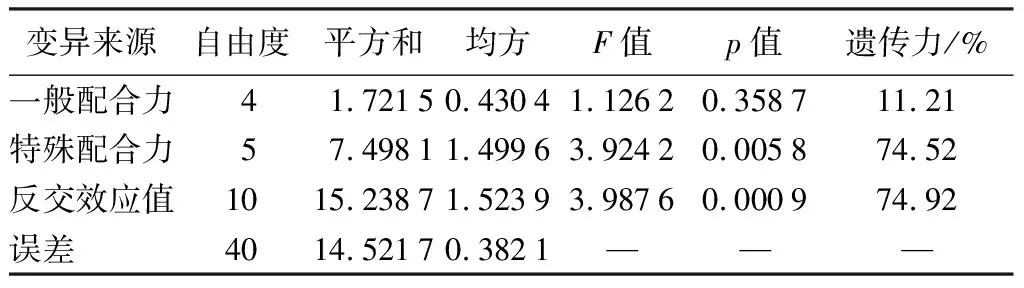
表6 白桦双列杂交亲本碳储量配合力及反交效应值的方差分析
对白桦各杂交组合的特殊配合力分析(表7)表明:单株碳储量的特殊配合力效应值均为正的组合为5×8(8×5)、14×5(5×14)、14×8(8×14)、和11×15(15×11),效应值分别为0.378、0.091 2、0.799、1.268 3,说明这些组合的杂交子代均优于双亲的平均值,但是,由于经过母本父本间生长性状的差异显著性分析,5号亲本不适宜作为杂交父本(表7),不宜选择8×5,14×5,但5×8和5×14是5号做母本的优良组合。因此,选择14×8(8×14)、11×15(15×11)以及5×8和5×14为待选的高碳汇优良组合。单独分析特殊配合力只表明了该组合的子代表现优于双亲的平均表现,但是考虑到父本母本的选配时,应使用反交效应来分析,并且反交效应值的方差分析(表8)表明其达到了的极显著水平,说明亲本之间的选配也存在着显著的差异。
对上述入选的各组优良杂交组合的反交效应值分析(表7),适宜的优良杂交组合为8×14、15×11、5×8和5×14,对应的家系为619、643、602、605,再对应单株碳储量多重比较,602、619和643家系包含在初步待定的单株碳储量平均值较高的6个家系里,因此确定为亲本配合力较好的高碳汇家系。
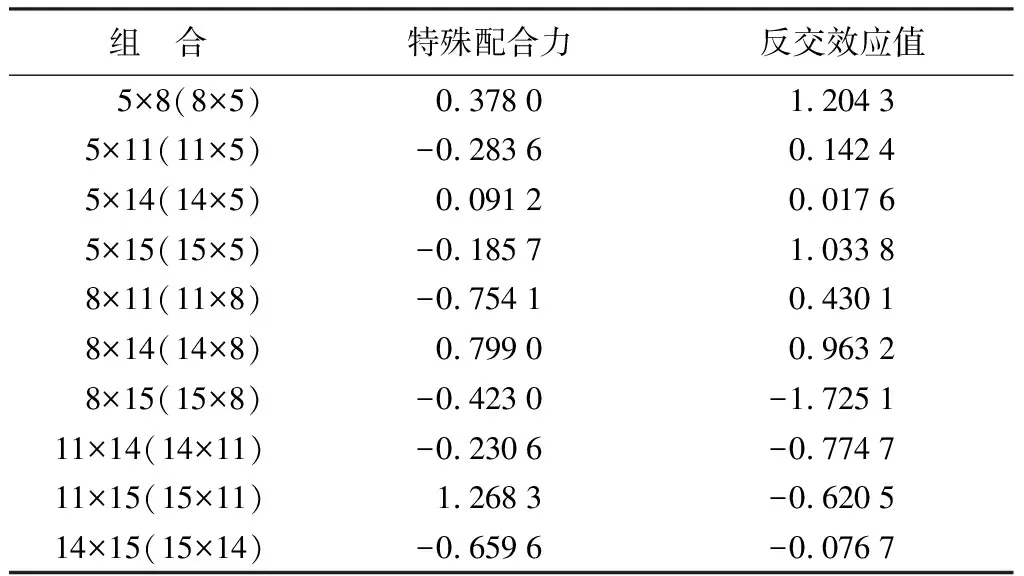
表7 白桦各亲本组合碳储量特殊配合力及反交效应值
4 结论与讨论
应用双列杂交设计控制杂交,得到全同胞子代进行高碳汇家系及亲本杂交组合的筛选,选配出高质量并且具有高碳汇能力的家系及优良亲本,可以根据固碳要求提高育种成效,也为生产造林和生态系统碳平衡的紧密联系提供理论依据。但是,对碳储量的研究方面不能仅限于对子代的筛选,还需对杂交亲本进行选配。林木育种研究者常根据配合力的大小选配杂交亲本[16,24-26]。本研究通过对白桦5×5双列杂交试验获得的全同胞子代测定林生长性状和碳含量调查研究,初步筛选出627、637、602、642、643和619家系单株碳储量较各家系平均值高13.69%~43.67%,确定为高碳汇优良家系,并且对其各自的杂交亲本作为优良杂交亲本继续进行亲本分析。对各杂交组合母本、父本的配合力和反交效应值进行分析发现,杂交亲本的一般配合力差异不显著,而特殊配合力和反交效应值均表现出了极显著的差异,说明在众多杂交组合中,同一亲本作为父本或者母本,家系间变异较大。利用一般配合力高的亲本杂交可望获得好的杂交效果,而特殊配合力高则是选取优良杂交组合的依据之一[12]。因此,筛选特殊配合力高的杂交组合意义重大。对入选杂交组合的特殊配合力和反交效应值进行计算,发现有些组合的杂交子代的特殊配合力均优于双亲的平均值,对这些组合通过反交效应再进行筛选,8×14、15×11、5×8和5×14的特殊配合力较高,再对应其子代的单株碳储量分析,确定优良碳汇杂交组合为8×14、15×11、5×8。
由于碳储量由生物量与碳质量分数乘积得到,碳储量受生物量和碳质量分数共同影响。单株碳储量的显著差异主要是由地上部分碳储量的显著差异所影响,而地上部分碳储量的显著差异是由树干、树叶、树枝的碳储量显著差异造成。再根据变异系数分析,地下部分以及树根的碳储量变异系数明显较大,且地下部分的生物量无显著差异,因此设想可能是由于变异大,更容易造成其家系间差异的不显著。在筛选优良家系的过程中,尽可能排除变异系数较大的家系或亲本,以保证其遗传稳定性。树皮的生物量和碳含量以及碳储量均无明显差异,并且碳储量很低,推测原因是树皮在固碳机制中的作用很小;而树干则是参与碳储存和运输的重要部位,其碳质量分数差异极显著,生物量和碳储量也在0.05水平上差异显著,因此树干在碳汇方面是需要考虑的重要因素之一;叶作为CO2固定的初始部位,在三者中都表现出极显著的差异,因此在固碳能力方面应被着重考虑;枝的生物量差异显著,与其生长及分枝情况有关,但其碳质量分数无明显差异,猜测其碳储量的差异主要由生物量导致,应对其再做进一步研究;根部碳储量的差异不显著主要是由生物量差异不显著所影响,推测其碳含量的显著差异与根呼吸释放CO2的机制有一定关联,可以将根呼吸作为碳释放因子在研究工作中着重考虑。
亲本的配合力并不是指其本身的表现,而是指与其他亲本结合后它在杂种世代中体现的相对作用。在杂种优势利用中,配合力常以杂种一代的产量表现作为度量的依据;在杂交育种中,体现在杂种的各个世代,尤其是后期世代。因此,通过双列交配设计来估算亲本配合力和反交效应,是杂交育种亲本选配的必要条件。一般配合力效应值高的亲本也较多表现出特殊配合力效应值较高,但也有例外,表现为一般配合力低的亲本和一般配合力高的亲本杂交也会产生高的特殊配合力,14号杂交亲本是从芬兰引进的欧洲白桦优树,具有干型通直、树形美观等特点,但是其抗寒性较差,而与其杂交的中国白桦优树,抗寒性强,其杂交子代继承了亲本的优点,干型通直、且抗寒性强,因此选择欧洲白桦作为杂交亲本具有重要的潜在价值[13]。
本研究筛选获得的高碳汇家系可以进行培育,获得其无性系子代可投入生产造林和营建种子园,以保存其亲本的高配合力,保证优良性状和高碳汇能力的稳定遗传,从而为生态系统的碳平衡的起到积极作用。对于其无性系子代,需进行繁殖和培育,并且进行光合和呼吸作用与固碳机制方面的深入研究。未来的遗传育种工作中,在将干型、材质材性、适应性等作为选择育种依据的同时,不妨把固碳能力也考虑在内,将育种与生态系统平衡紧密联系起来,以获得多方面的收益。
