城市绿化植物群落结构分布格局与多样性分析
2018-08-04庄华蓉
庄华蓉
(重庆城市管理职业学院,重庆 401331)
【研究意义】物种多样性,又称群落多样性或生态多样性,具体是指物种水平上表现为多种生物的形式,学界对于生态层面上物种分布的均匀程度主要是借助对群落组织水平的研究来实现的[1-2]。通常而言,在研究生物多样性过程中,对于其组成和结构上多样性的研究是对其组织水平和功能状态进行科学认识的最重要的基础[3-4]。由此可见,研究群落多样性不仅能够了解物种空间分布规律,而且能够科学的解释群落与环境之间的相关关系。“绿化植物是由城市区域内所有树木所组成”,对于城市而言,其园林绿地系统的主要组成部分就是绿化植物。对于群落结构的研究有许多方面,通常是从以下方面着手:种群动态与结构、物种组成、空间布局、层片结构等人口高度密集的城市环境特点[5]。群落结构能够有效的改善城市环境,保障城市景观丰富、绿地健康和稳定,是一个城市生态的基础[6-7]。植物多样性能够体现一个城市的绿化植物群落是否稳定和健康,即植物的丰富度、多样性、均匀性等特点能够直接反映城市群落的各项特征,比如结构、组织化水平、功能等[8]。【前人研究进展】研究生物多样性的关键主要体现在物种组成及多样性,而不同群落多样性差异明显,一般而言,主要体现在群落组织水平、结构类型以及群落之间的稳定程度。生态系统的一个基本特征就是物种多样性,其决定了整个生态同的功能和稳定。因此,衡量生态系统管理、生物多样性保护、生态系统各个功能方面的一个重要指标就是物种多样性[1-2]。最近几年来,森林公园、自然保护区、城市绿地土壤养分是研究的重点,但是在研究城市绿化植物群落多样性方面略显不足[9-11]。作为我国一个典型的山地城市,重庆市有着起伏不平的地形,使得城区各地高差不一,不利于城乡交错、连片发展。穿城而过的长江及支流嘉陵江使重庆城区的布局较为分散,进而造成了其山城独有的各种植被种类组成和分布的特色[12-14]。【本研究切入点】近10多年来,我国城市建设和城镇化的不断加快,城市不断向外扩张的行为使得周边环境、林地、生物多样性都受到严重损害。在研究城市至郊区之类梯度变化方面,国外学者起步较早,随着我国城市化建筑随之产生的环境问题日益严重,我国学者对于诸如城乡绿地空间梯度变化等城市至郊区植被梯度变化的研究不断受到人们的重视[15-17]。此外,我国对于植物群落的环境梯度变化的研究也不多,尤其是通过对各群落类型的物种多样性对比分析,并结合城市绿化植物重要值来对优势群落类型进行分析的相关研究更是少之又少。【拟解决的关键问题】基于此背景,本研究力求在深入调查和分析重庆市城区至郊区植被梯度变化现状的基础上,对重庆市植被生态梯度组成及特点特的研究,旨在为合理规划城市植物、科学保护城市植物多样性、科学调整城市园林植物应用提供科学的基础数据。
1 材料与方法
1.1 研究区概况
重庆市地理位置独特,东面、南面、北面都是山区,整个城市被江河和山体分成不同的独立区块。平坝、丘陵、低山和中山是其地貌的具体特征,由此可见重庆市有着较为复杂的地形地貌结构。重庆市整体面积为8.2万 km2,其中18.2 %为丘陵,75.3 %为山地,其主城区是一个半岛形状,三面环水,地处嘉陵江与长江交汇的中亚热带湿润季风气候的丘陵地带,其降水量非常充沛,常年高达1000~14 000 mm。年平均日照时数和年平均太阳辐射量较高,尤其是在夏季,高温日数较多,天气炎热。
1.2 样地设置
通过选取重庆主城区四条宽度一样,长度不同,且不同走向的样带进行研究(表1),分别是“渝中区-南山-明月山脉”、“沙坪坝-歌乐山-虎峰山”、“江北区-复兴镇”、“九龙坡区-桥口坝风景区”,长度分别为20.6、25.3、20.4、25.1 km,走势分别为自西向东、自东向西、自南向北、自北向南。在地形上,样带1和样带2存在着较大的起伏,这就导致不能在主城区进行连片、集中的发展建设用地,因为存在着较大的相对高差,也造成了主城区之间存在的模式是“城市-山脉-城市-山脉”;在地势上,样带3和样带4则表现的较为平坦,但该区域主要是农田为主,具体模式为“城市-耕地”。
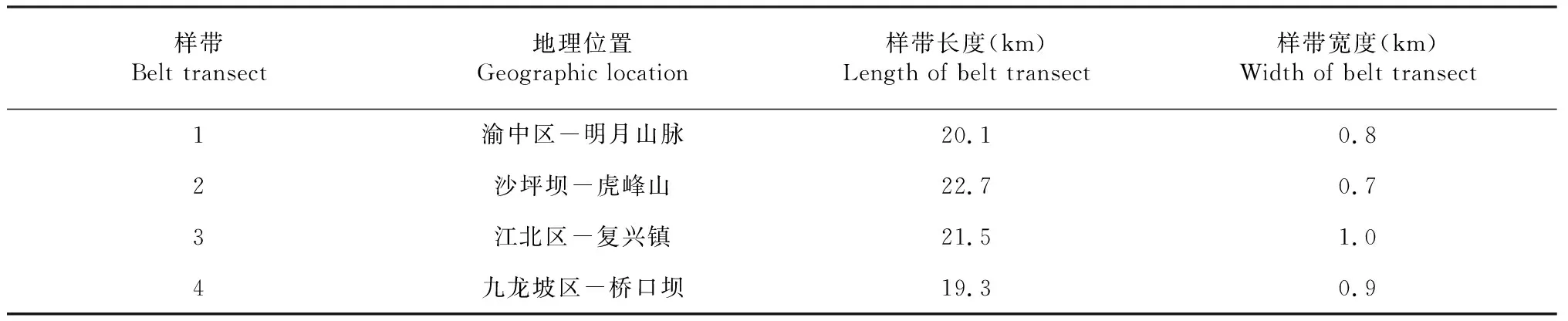
表1 样带概况
1.3 数据分析
在本研究中,样带1至样带4取样起点都是重庆市区内繁华的步行街,分别为解放碑步行街、沙坪坝三峡广场、观音桥步行街、杨家坪步行街,这样选择的目的是为了更好的对城市至郊区植被的梯度变化规律进行分析。对于样地的设置方面,首先要结合重庆市整个城市的特点,然后在科学参考国外《“城-郊-乡”森林生态样带植被变化梯度分析》的基础上来进行设定,所以本研究采用的是1 km×1 km样地设置方法。步骤是在重庆市区内选好的4个样带中分别选择2个1 km×1 km的样地。选好之后再每隔2~3 km取1个1 km×1 km的样地。然后再跟《土地利用现状分类》国家标准GB/T21010-2007中对于用地类型的规定和要求进行样方的选择,即在建设用地、公园农业用地、森林、灌草以及裸地中科学的选择数量约为10~20个的10 m×10 m的样方。选择完成之后,还必须明确和掌握各个样方的位置特征,同时详细的记录各个样方区域内各个木本植物的数量和种类。
α多样性指数选取Simpson优势度指数、Shannon-Wiener多样性指数、Margalef丰富度指数、McIntosh均匀度指数[1-3]:
Shannon-Wiener指数(H):H=-∑(PilnPi)
Margalef指数(M):S=(N-1)/lnN
Simpson指数(D):D=1-∑(Pi)2
N表示植物总数,ni表示第i种的个体数量,Pi表示ni/N。
Cody指数和Sorenson指数表示β多样性[1-3]:
Cody指数(βc)=(G+L)/2
Sorenson指数(Cs)=2j/(a+b)
式中:G代表沿着生态梯度增加的物种数目;L代表沿着生态梯度减少的物种数目;j代表共有的物种数目;a和b分别代表样地A和样地B的物种数目。
Excel 2010.0和SPSS 21.0进行数据统计和方差检验,以平均值±标准误差表示(Mean±SE),单因素方差分析(One-way ANOVA),CANOCO 5.0分析环境因子对植被多样性的响应。
2 结果与分析
2.1 样地基本特征
由表2可知,4条样带的主要用地类型均为建设用地,受人为干扰的影响最大。郊区则以森林、农业用地和灌草为主,受人类干扰程度较小。由样带1到样带4,受干扰程度依次降低。
2.2 乡土树种比例
乡土植物指的是在某些特定地区,与当地土壤、气候等自然条件达到一定的平衡,并稳定、自然分布,且不受人力干扰作用的树种。在本研究中,研究所需的乡土树种的确定标准主要是依据《重庆维管植物检索表》。本研究中所选择的4条样带中,木本植物物种中城区乡土物种所占的比例分别为,样带1为42.2 %、样带2为46.7 %、样带3为35.4 %、样带4为25.9 %。原生乡土树种比例远远小于外来物种说明人化工和高度城市化影响了重庆市的城市园林植被。由于样带1至样带4在位置、地形等方面存在差异,之间的木本植物物种中近郊乡土树种的比例自然存在不同,比例最高的是样带2,为72.1 %,其次是样带1占比65.7 %、样带3和样带4最小,分别占比为59.3 %和52.7 %,木本植物物种中远郊乡土树种的比例从最高到最低分别为样带2占比70.2 %,样带1占比69.8 %、样带3占比64.3 %、样带4占比58.5 %,这主要是因为在城区植被受到人类活动和行为干扰程度大于郊区植被,导致了整个重庆市郊区木本植物中超过一半以上的都是乡土物种(图1)。

表2 各样地基本特征
不同小写字母表示差异显著(P<0.05),下同Different lowercase letters indicate significant difference (P<0.05), the same as below图1 城市乡土树种比例Fig.1 Proportion of urban native species
2.3 植物α多样性变化
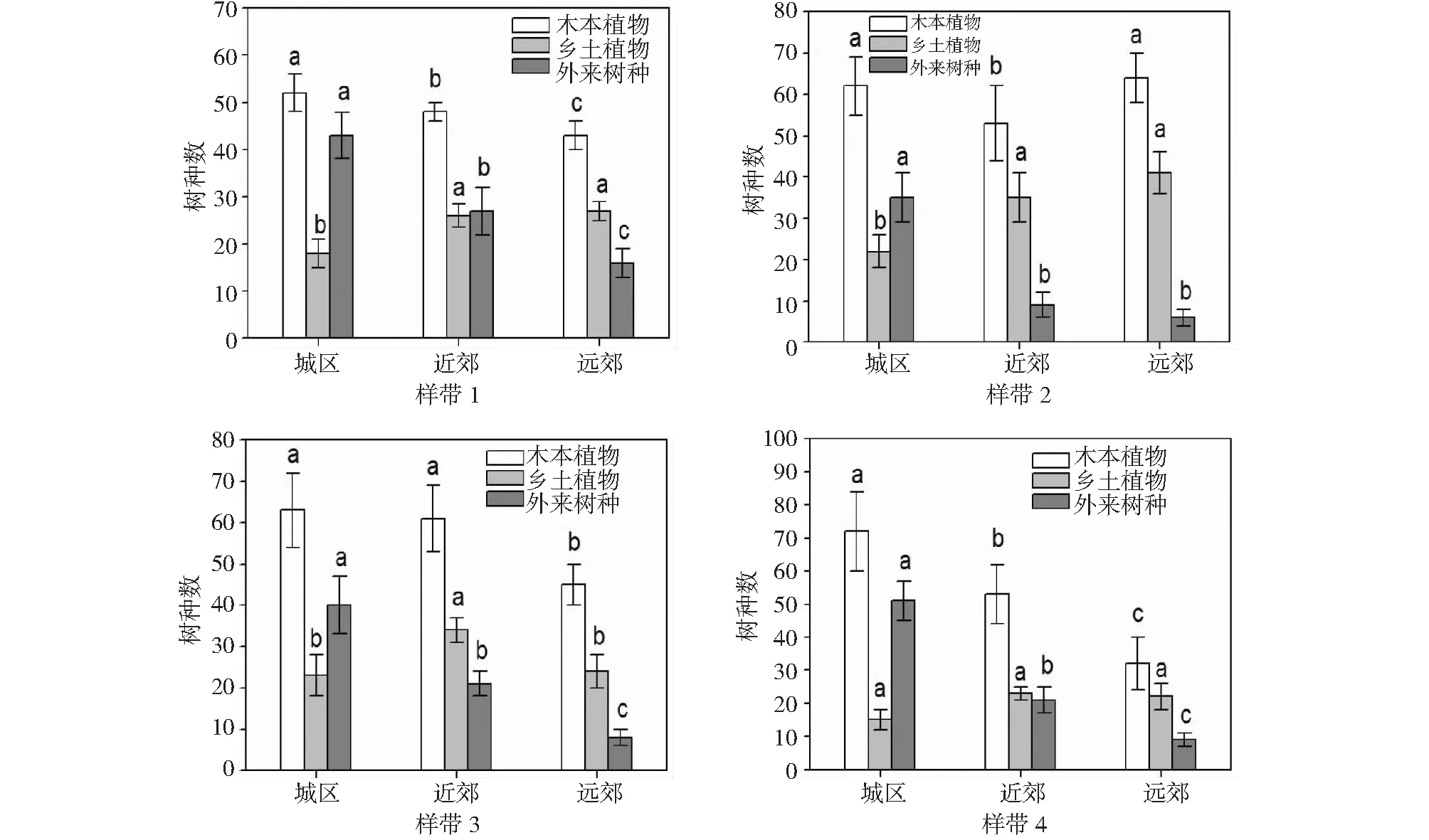
2.3.1 Margalef丰富度指数 不管是城市的远郊还是近郊,其在物种丰富度方面,远比不上城区,因为城区存在普遍较大的物种丰富度。这也归功于最近几年来重庆市城市绿化措施的提升,同时对于绿化建设方面的重视也日渐加强,即在对于多种类树木引进的同时更加注重其物种多样。本研究中选取的样带1和样带2为“城市-山脉-城市-山脉”模式,从远郊到近郊呈现逐渐减小趋势是其物种丰富度的特点,样带3和样带4则为“城市-耕地”模式,其物种丰富度的呈现特点则与“城市-山脉-城市-山脉”模式相反,这主要是地形因素和人为因素引起。对于样带1和样带2而言。因中梁山和铜锣山自北向南的贯穿而使得其用地类型、地形变化等方面都存在很大的差异而导致一定的复杂性。但是样带3和样带4不管是从城区到远郊的地形上,还是在用地类型方面,都会比样带1和样带2简单许多(图2)。
2.3.2 Shannon-Wiener多样性指数 由图3表明,样带1、样带2、样带3、样带4木本植物不管是在远郊、近郊还是城区,都存在着较大的物种多样性指数,最为突出的就是在城区,其物种多样性指数高达0.95以上,在远郊及近郊则稍微小一点,总体而言,就物种多样性指数而言,不同的样带呈现出来的特点自然不同,本研究中所选的4条样带也是如此,多样性指数最高的是样带1、样带2、样带3、样带4的城区,其次是样带1和样带2的远郊和样带3、样带4的近郊,最低是样带1、样带2的近郊和样带3、样带4的远郊。这个特点与木本植物在样带1至样带4物种丰富度指数规律方面基本相同。
2.3.3 Mclntosh均匀度指数 由图4表明,在均匀度指数方面,4条样带都表现出城区均匀度指数较大的特征,最小的地方为近郊。4条样带木本植物均匀度指数存在差异,其中样带1与其他3条样带不一样,远郊的均匀度指数最大,其次是城区,最小的近郊,而样带2、样带3、样带4则不然,均匀度指数最大的为城区,其次是远郊、近郊。由于人为的干扰,郊区木本植物群落在均匀度指数方面不高,但是有较高的多样性指数,原因体现在以下两个方面,一主要是因为像是次生竹丛、香樟、喜树等这样的乔木在达到一定的高度和树龄之后对人为干扰及其他行为敏感度较低;二是植物生境因为人为的干扰而产生的变化导致了部分乔木和灌木幼株的更新,使得木本植物群落均匀度下降,但多样性升高的情形。
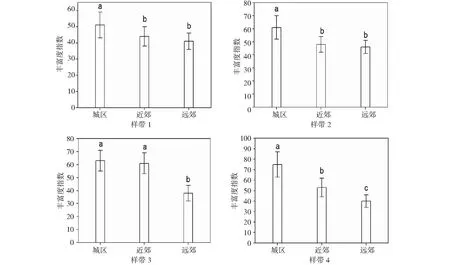
图2 城市绿化植物Margalef丰富度指数Fig.2 Margalef index of urban greening plant
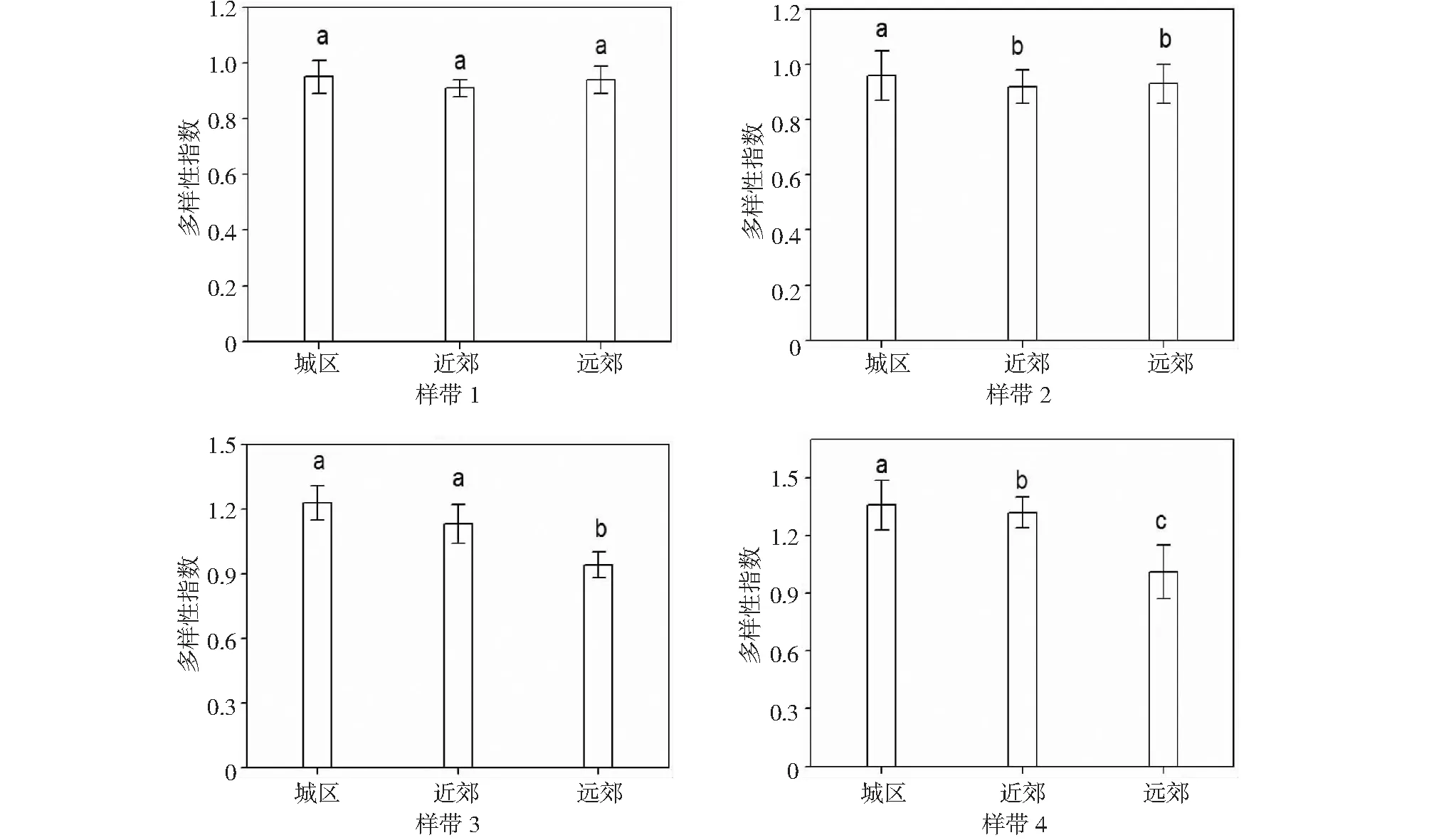
图3 Shannon-Wiener多样性指数Fig.3 Shannon-Wiener index of urban greening plant
2.4 植物β多样性
2.4.1 Cody指数 Cody指数在获取有关物种替代信息方面有着重要的作用,原理是比较新增加和失去物种数目的方式来获取。在真实反映群落功能和结果方面,β多样性是一个反映地区不同群落沿环境梯度上不同的物种进行更替的相关速率,β多样性越大就表明其在进行物种替代时的速率越高。反之,则替代速率就小。由图5显示,在Cody指数方面,4条样带表现一致,最高的是近郊,其次是远郊,最小的是城区。并且这4条样带不管是在城区、远郊、近郊上的带Cody指数差异都较为显著(P<0.05)。

图4 Mclntosh均匀度指数Fig.4 Mclntosh index of urban greening plant

图5 Cody指数Fig.5 Cody index of urban greening plant

图6 Sorenson指数Fig.6 Sorenson index of urban greening plant
2.4.2 Sorenson指数 由图6可知,4条样带的相似性系数均表现为远郊>近郊>城区,其中远郊和近郊Sorenson指数差异不显著(P>0.05),二者显著高于城区(P<0.05),由此表明,近郊和远郊区对的相似性指数较大,说明二者之间共有物种数较多;而近郊和城区资源异质性差异较为明显,说明二者之间的共有种数较少。
2.5 植物多样性排序分析
植物分布差异的主要是环境因子变化所致,外界环境能够较大的影响到植被多样性。当前,在研究环境因子与植被多样性之间复杂关系中,应用最为广泛的方法就是能够对环境因子和植被的相关关系进行客观反映的排序图,由于造成植物分布差异是因为环境因子,所以在本研究中的相应变量就设定为植被丰富度指数,优势度指数、均匀度指数以及多样性指数,解释变量则为环境因子,这样做的目的是更好的对环境因子与植被多样性之间存在的相关关系进行科学的揭示。在研究过程中,为了对能够明显解释影响植被变化的指标进行准确的提取,采用的方法是多元统计分析方法。PCA排序图优点众多,其中一个是保证在独立保持各变量对环境的贡献率前提下来描述单个变量的特征,这种特征描述是建立在对各个变量进行不同组合的形式上,同时在一个图上同时来排序研究对象和环境因子(图7和表3)。图7显示,箭头所指的象限表示排序轴与环境因子之间存在的相关关系,且这种相关关系与箭头的长度相关,相关性越大,则箭头连线的长度自然也就会越长。与此同时,箭头的夹角也与相关相关程度紧密相连。此外,还与两个箭头之间以及排序轴与之的夹角大小相关,如果两者之间的夹角大于90度,则这两者的相关关系为负相关。在PCA排序图中,0.789与0.124为前2个排序轴特征值。由图7可知,反映梯度变化指标特征的是第一排序轴,所谓的梯度具体是体现在环境因子与不同植被多样性之间,两个排序轴之间存在着相关性都为1.000的显著相关关系。除此以外,在对于变量的解释率方面,两个排序轴都高达99.13 %,在对植被多样性受环境因子影响分析过程中,通过利用蒙特卡罗检验可以发现,影响非常显著(第一轴P<0.000,F= 8.69;第二轴P<0.000,F= 6.15),由此可见,在解释环境因子对植被多样性的影响方面,PCA排序图作用明显。相关研究显示优势度指数、多样性指数、丰富度指数之间都存在着显著的正相关关系,同时也和土壤各养分之间存在着显著的正相关关系。在PCA排序轴的第一排序轴中,均匀度指数、多样性指数以及丰富度指数与显著性影响因子(环境因子)之间存在着显著的正相关关系,即随着环境因子的增加而增加。综上所述,土壤养分是影响绿化植物α和β多样性指数的变化的主要因素,其中影响比较显著的有全氮、有机碳、土壤含水量的大小。此外,对于植物多样化指数,温度、电导率、土壤pH值的也会起到一定的抑制作用。
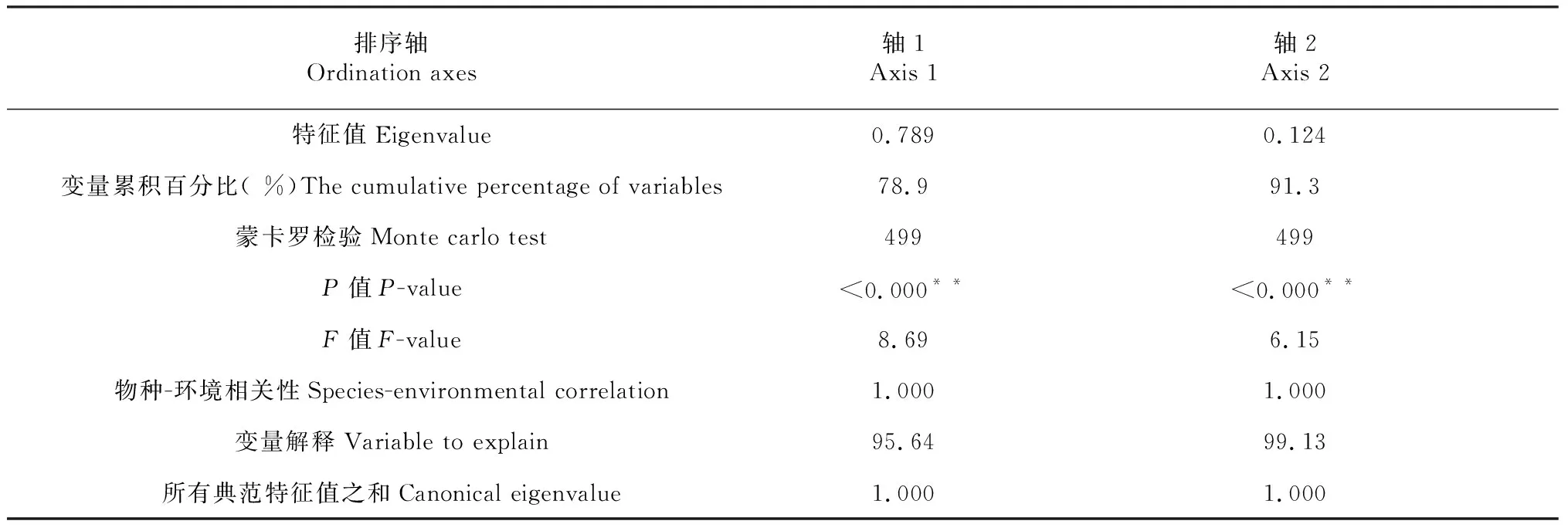
表3 植物多样性排序
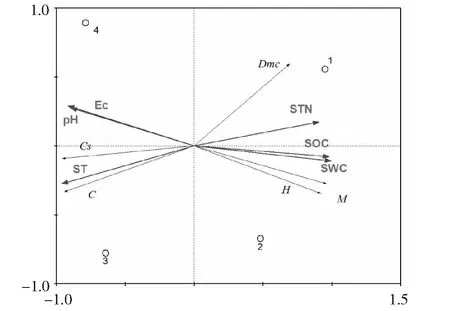
M-Margalef index;H-Shannon-Wiener index;D-Simposon index;Dmc-McIntosh index;C-Cody index;Cs-Sorenson indexEc-电导率(Electrical conductivity);SWC-土壤含水量(Soil water content);ST-土壤温度(Soil temperature);SOC-土壤有机碳(Soil organic carbon);STN-土壤全氮(Soil total nitrogen)图7 植物多样性排序Fig.7 The plant diversity ordination
3 小 结
物种多样性主要反映在外界条件综合作用下的植物群落,物种多样性的内涵主要表现在单位样方物种丰富度、总种数和Simpson指数以及Shannon-wiener指数方面,总种数的大小也会受到物种丰富度(样方内物种数)以及Shannon-wiener指数(样方间物种周转速率)的影响[18-19]。Margalef指数和β多样性之间的相关性比较显著,且在4条样带中变化的规律都是一致的,由此可知,样方物种丰富度能够较大的影响到样地总种数,重庆市在植物群落组成方面一致性较强,在β多样性方面,也存在着较大的变异性,原因与环境相关的可能性较大[20-22]。
由此可知:在样带中,城区和远郊之间存在着明显的梯度,在物种多样性和丰富度方面,城区明显高于郊区,此外加之乡土树种比例的不同使得之间的植被变化明显。这得益于重庆市在最近几年对于物种多样性在城市园林绿化中的重要性越来越重视的缘故。在物种的多样性指数和物种的丰富度指数方面,远郊和近郊存在明显差异的原因是:从地形上来看,重庆城区比较复杂;从干扰上来看,人类活动较多;从用地类型来看,各个地块所占比例差异较大。城市化进程的加快,使得郊区也面临着各种城市建设问题,在此过程中必须注重自然植被的保留,如果存在无法保留的情形,则在郊区的城市化完成后利用生态恢复技术,借鉴相关群落特点来构建具有一定特色的地带性植物群落。
重庆市园林绿化的特点是从城区到远郊,木本植物中乡土树种的比例一直在提高,对于树种的应用存在不足是当前重庆市园林绿化建设的主要问题。人为活动干扰能够影响到木本植物中乡土树种的比例大小,干扰越小,所占比例越大。如何在充分挖掘自身植物资源的基础上,一方面注重城市物种多样性,另一方面凸显地域植被特征就成为重庆市园林绿化建设中亟需考虑的问题。应该保护郊区范围内的自然植被,同时在城市园林绿化建设中引入那些已经驯化的乡土树种[23-24]。能独立保持环境因子对不同群落变化的贡献率是PCA作为一种直接梯度分析方法的最大优势,此外,还能对其外部环境也呈现出显著的反馈作用。多样性对土壤养分反应均比较敏感,这一点主要体现在图7之中环境因子箭头、连线长度、夹角大小之上。敏感指标一方面是对其境指示作用的反映,另一方面也是对该区域内部的各个生态系统敏感性的一个重要表征。对于生态因子与植物多样性之间存在着何种内在联系,还需要进行深入的研究,只有这样才能对该区群落分布结构和格局的根本原因进行科学和准确的揭示。
4 结 论
(1)不同绿化植物样带城区木本植物Margalef丰富度指数均高于近郊和远郊区,依次表现为:城区>近郊>远郊。木本植物Shannon-Wiener多样性指数从大到小顺序依次为:城区、远郊、近郊;木本植物Mclntosh均匀度指数从大到小顺序依次为:远郊、城区、近郊;Cody指数均表现为近郊>远郊>城区。
(2)不同绿化植物样带Cody指数在近郊、远郊和城区差异均显著(P<0.05),相似性系数均表现为远郊>近郊>城区,其中远郊和近郊Sorenson指数差异不显著(P>0.05),二者显著高于城区(P<0.05)。
(3)PCA结果显示:绿化植物Margalef丰富度指数、Shannon-Wiener多样性指数、Mclntosh均匀度指数之间均呈正相关;沿着PCA的第1排序轴,随着显著性影响因子(含水量、有机碳、全氮)的增加而增加,其中土壤含水量、有机碳和全氮的影响为正;土壤pH值、电导率和温度在一定程度上抑制植物多样性指数的变化。
