谷子Hd1-like基因的克隆及其与农艺性状的关联分析
2018-08-02袁玺垒戴凌峰张小梅郁飞燕贾小平
袁玺垒,戴凌峰,张小梅,郁飞燕,贾小平
(河南科技大学 农学院, 河南 洛阳 471023)
CO(CONSTANS)基因是在拟南芥中克隆的光周期途径中的关键基因,编码含有2个锌指结构的蛋白质,位于N末端的B-box结构域参与调节蛋白质之间的相互作用,而且在C末端含有核定位所需要的CCT域[1]。在长日照条件下CO基因通过促进FT基因表达而使开花提前。Hd1是在水稻中克隆的拟南芥CO基因的同源基因,位于第6号染色体,但是其作用方式却与CO基因不同。Hd1在水稻开花调控途径中具有双重功能,在短日照条件下促进开花,而长日照条件下抑制开花[2-3]。在对其他植物CO基因相关研究中发现,无论单子叶植物还是双子叶植物,其CO同源基因都可以互补拟南芥CO基因突变产生的晚花表型,说明单子叶植物和双子叶植物CO基因在调控成花方面具有保守性[4]。研究表明,Hd1基因是一个多效基因,除影响作物抽穗期外,还影响株高、穗粒数等多个农艺性状。如Hd1基因和Ehd1基因可以减少水稻穗部初级分支数目,使每个穗的小穗数目减少[5];构建遗传背景一致的2个BC2F5群体对水稻RFT1-Hd1a所在区间、Hd1所在区间进行QTL分析,发现这2个区域都对抽穗期、株高、千粒质量有显著效应,但是Hd1所在区间作用明显强于RFT1-Hd1a所在区间[6];以珍汕97为轮回亲本、密阳46为供体亲本的BC2回交组合,构建世代分别为BC2F6和BC2F7的NIL-F2群体和世代为BC2F8的近等基因系群体,这些群体的QTL分析结果表明,在珍汕97遗传背景下,来自珍汕97的Hd1感光等位基因缺失了光周期敏感性,在自然短日照和长日照下均促进抽穗,并同时降低株高、粒数和产量[7]。
谷子(SetariaitalicaBeauv.)属于短日照喜温作物,对光周期和温度反应比较敏感是导致其生态适应性狭窄、生产上缺少跨大区种植当家品种的重要原因。其中,控制抽穗期的基因对谷子生态适应性有重要作用。目前,谷子全基因组序列已经测定,分析表明,谷子抽穗期主要依赖光周期,其中CO基因和Ehd1、Ghd1基因是控制抽穗期的关键基因[8-9]。而谷子中水稻Hd1同源基因的相关研究报道极少,近期有学者通过对高粱、水稻、谷子进行比较基因组学研究,发现在谷子4号染色体存在水稻Hd1基因的同源基因,关联分析表明,谷子Hd1中存在一个高频的从GT到AT的剪接位点变异,该变异导致谷子的生育期变长,Hd1基因在水稻、高粱、谷子中受平行驯化[10]。这是唯一有关谷子Hd1基因的报道,但是该研究没有阐明谷子Hd1基因是否具有多效性。本研究利用谷子基因组序列信息从11个谷子品种中克隆水稻Hd1基因的同源基因,并且分析其在不同谷子品种间的变异情况,揭示Hd1基因变异与谷子各农艺性状的关系,明确该基因是否具有多效性,同时也为进一步揭示谷子的光周期反应分子机制奠定基础。
1 材料和方法
1.1 植物材料
11个谷子品种为豫谷1号、郑谷2号、郑8041(14)、济叶冲4、冀谷1号、安04-5014、龙谷26号、郑州12、8322-14、法谷28-81、冀谷27。
1.2 农艺性状调查及分析
谷子材料于2014年种植于河南科技大学周山校区试验田,种植方式:每个品种2行,行距30 cm,株距5 cm,行长2 m。于谷子生长期调查抽穗日数,抽穗日数=抽穗期-出苗期;株高、穗长、穗粗、穗质量、码粒数、穗码数、穗粒质量、千粒质量,每个品种选中部10株于成熟期及采收后进行性状测量,性状测定方法参照文献[11-13]。用SPSS 18.0软件进行品种间各性状差异显著性检验及性状间相关性分析。
1.3 谷子Hd1-like基因的克隆
采集谷子幼嫩叶片,用常规的CTAB法提取11个谷子品种基因组DNA,从phytozome 10.0数据库中获得谷子Hd1-like基因序列(GenBank登录号:AB807721.1),用DNAMAN 5.0软件设计一对特异引物用来扩增包含所有编码区、大小为2 340 bp左右的谷子Hd1-like基因序列。引物序列为:正向5′-TGCAAGTGCCAAGCTAAG-3′,反向5′-CCAAGTTCCTTCAAAACCAT-3′。扩增体系25 μL:DNA模板50 ng,引物2.5 pmol/L,扩增 MIX12.5 μL,ddH2O9.5 μL。扩增程序:94 ℃预变性5 min;94 ℃变性40 s、51 ℃退火1 min、72 ℃延伸1.5 min,共35个循环;最后72 ℃延伸10 min。
1.4 基因突变位点检测及突变位点与农艺性状关联分析
对获得的Hd1-like基因序列使用多序列比对软件ClustalX进行多序列比对,寻找突变位点;用TASSEL 2.1软件进行LD分析及表型性状与多态性位点的关联分析。
2 结果与分析
2.1 谷子品种间各性状差异显著性分析及性状间相关性分析
对包括抽穗日数在内的9个性状进行相关性分析,结果表明,抽穗日数与株高、穗长、穗粗、穗质量、穗粒质量存在显著正相关(P<0.05),与码粒数存在极显著正相关(P<0.01);株高与穗码数、千粒质量存在显著正相关(P<0.05),与其他各穗部性状存在极显著正相关(P<0.01);穗长与其他各穗部性状间存在显著或极显著正相关(表1)。

表1 谷子性状间的相关性分析
注: *表示在0.05水平(双侧)上显著相关; **表示在0.01水平(双侧)上显著相关。
差异显著性分析结果表明,株高、穗长、穗粗、穗质量、穗码数、码粒数、穗粒质量、千粒质量8个农艺性状在11个谷子品种间均存在显著差异,多数性状在品种间存在丰富的变异类型。株高存在极高类型如郑8041(14)、郑州12、8322-14、法谷28-81,株高均超过127 cm;较高类型如豫谷1号、郑谷2号、安04-5014,株高在115~125 cm;中间高度类型如济叶冲4和冀谷1号,株高在107~113 cm;矮秆品种如龙谷26号和冀谷27号,株高小于70 cm。穗长也存在极长型(大于20 cm)、较长型(15~20 cm)、中间型(10~15 cm)、较短型(小于10 cm)4种类型。其他性状如穗质量、码粒数、穗粒质量也存在多种变异类型,表明本研究所选的11个谷子品种具有一定代表性(表2)。

表2 8个性状在11个谷子品种间的差异显著性检验
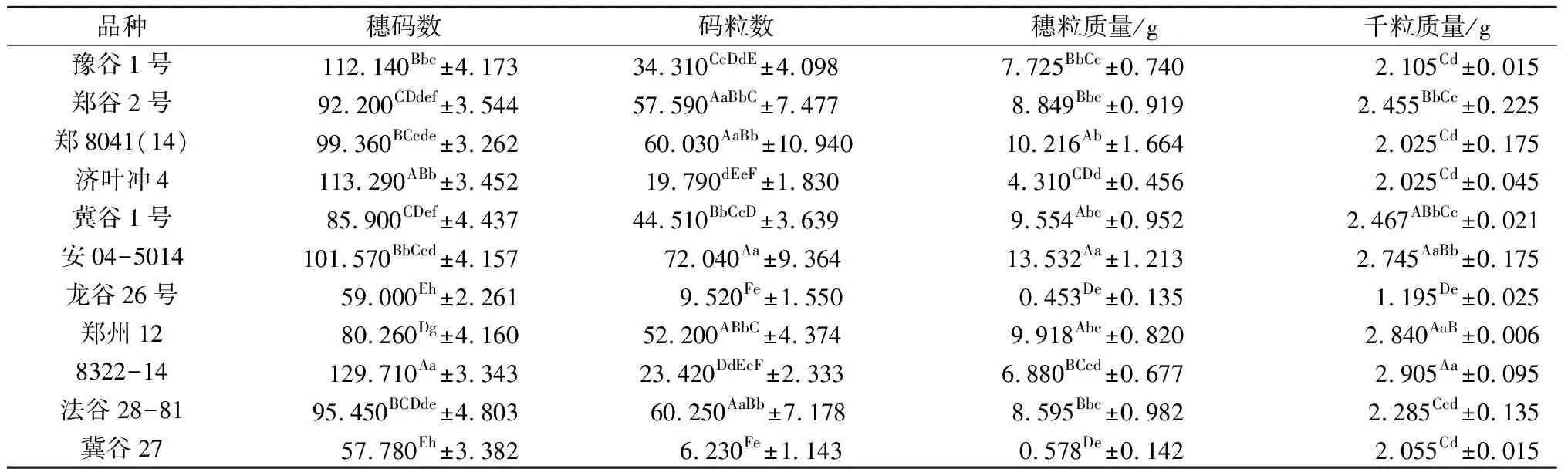
续表2 8个性状在11个谷子品种间的差异显著性检验
注: 同列均值右上角大写字母不同,表示差异极显著(P<0.01), 同列均值右上角小写字母不同,表示差异显著(P<0.05)。
2.2 谷子Hd1-like基因的克隆及序列分析
利用所设计的特异引物从11个谷子品种中均扩增出目标基因片段,大小与预期相符(图1)。扩增产物经克隆、测序,11个品种获得总长为2 351 bp的基因序列。对11条获得的Hd1-like基因序列进行多序列比对分析,在包含整个编码区、内含子区内共检测到43个突变位点,这些突变位点均为SNP,没有发现Indel。在编码区检测到24个SNP,内含子区检测到19个SNP(表3)。对检测到的SNP进行分析,发现核苷酸突变类型及它们的发生频率有所不同,主要为碱基A/G、T/C 2种替换类型(表3)。
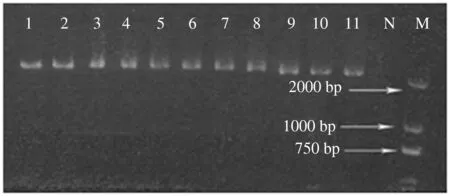
N:空白对照; M:DL2000 DNA Marker;1—11:分别表示供试11个谷子品种图1 11个谷子品种PCR产物电泳检测结果

表3 Hd1-like基因编码区检测到的SNP位点
注: 带阴影的位点位于外显子区,不带阴影的位点位于内含子区。
对43个SNP位点进行连锁不平衡(LD)分析,发现2个较大的连锁不平衡结构,均由6个位点组成,第一组包括位点71、1023、1117、1406、1875、2073,只有位点71、2073位于外显子区,其余4个位点均位于内含子区域;第二组包括134、185、921、1131、1187、2098,其中位点134和185在外显子区,其余4个位点在内含子区(图2)。

图2 Hd1-like基因突变位点间的连锁不平衡分析
根据基因组数据库phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)中提交的豫谷1号Hd1-like基因CDS序列(Seita.4G122700.1),对11个谷子品种Hd1-like基因推定的蛋白质序列进行了预测,结果发现,谷子Hd1蛋白和水稻、大麦、小麦等植物同源蛋白具有相似的结构特征,在靠近N端具有典型的锌指结构,靠近C端具有典型的CCT域(图3)。对11个推定的Hd1蛋白序列进行多序列比对,共发现15个氨基酸替换位点,这15个突变位点有2个位于保守的功能区域,其中第44个氨基酸突变位于锌指结构区域,该处氨基酸在10个谷子品种中为酪氨酸,而在郑州12中则突变为半胱氨酸。另一个突变位点位于CCT域,该区域第334个氨基酸在10个谷子品种中为精氨酸,而在郑8041(14)中突变为甘氨酸(图3)。由于这2个突变均为错义突变,因此可能造成郑州12和郑8041(14)两个品种对光周期敏感度减弱。

图3 11个谷子品种间Hd1-like基因推定蛋白质序列变异分析
2.3 谷子Hd1-like基因突变位点与主要农艺性状的关联分析
关联分析发现,有7个多态性位点与千粒质量存在显著相关,分别为:71、1023、1117、1406、1875、2073、2084,这些位点只有位点71位于编码区,剩下的6个位点均位于内含子区,除位点2084外,前面的6个位点均处在一个连锁不平衡结构中;有一个多态性位点与抽穗日数显著相关,为1165,该位点位于内含子中;与株高相关的多态性位点有2个,分别为1288和2270,其中1288位于内含子中,2270位于编码区;与穗粗相关的多态性位点有2个,分别为787和1938,均位于内含子中(表4)。
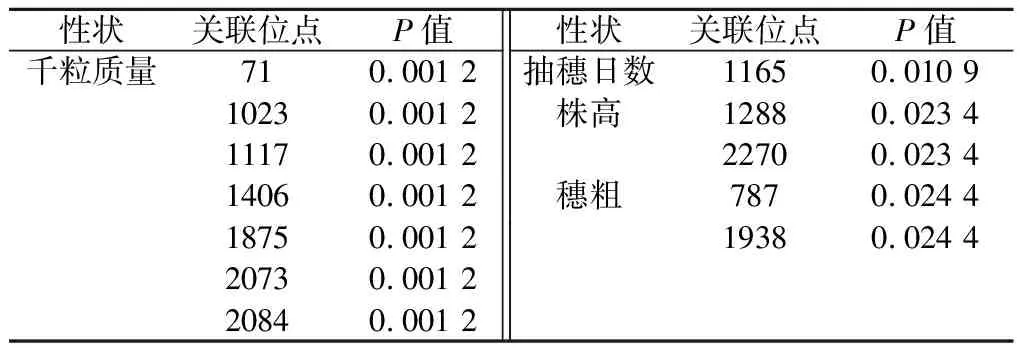
表4 与谷子表型性状关联的突变位点
3 结论与讨论
CO基因是植物光周期调控途径的关键基因,在长日照条件下促进拟南芥开花,而水稻中克隆的CO同源基因Hd1则在长日照条件下通过抑制Hd3a基因的表达而延迟开花。在高粱中,CO同源基因sbCO在缺失sbGhd7和sbPRR37的背景下无论长日照还是短日照均能促进开花[14]。因此,尽管在进化上具有保守性,不同植物之间CO基因的调控机制存在一定差异。本研究发现,谷子Hd1基因编码区产生的突变类型均为SNP,而无缺失/插入(Indel)类变异。而在水稻的研究中,发现Hd1基因编码区存在的3个插入突变和2个删除突变与开花时间高度相关[15]。在对高粱和谷子的研究中发现,Hd1基因编码区均存在插入/缺失突变,高粱Hd1基因编码区1个5 bp的删除导致移码突变,使抽穗期延迟,谷子Hd1基因共存在5处插入/缺失突变,包括1处6 bp的插入/缺失和4处单个碱基的插入/缺失,但是这些突变与抽穗期无关联,1个G/A替换导致的可变剪切与抽穗期紧密关联[10]。造成本研究未发现插入/缺失突变的主要原因是谷子材料的选择,本研究选择的11个谷子品种全部为栽培种,无野生种,而在Liu等[10]的研究中,选择的材料除栽培品种外,还包含15个野生品种,分析5处插入/缺失突变发现,有4处是由于野生种的引入产生的,只有1处单碱基插入突变是栽培品种产生的,因此可以推测,这4处插入/缺失很可能是在野生狗尾草向栽培谷子驯化过程中产生的,它们对野生种向栽培种的转变过程中可能起着某种作用。而1处由栽培品种引起的单碱基插入/缺失在本研究中未被检测到很可能是样本量的不同造成的,因为Liu等[10]的研究包含了80多个栽培品种,应该代表更大多样性,能够检出更多的突变位点。尽管本研究选择的材料有限,只有11个品种,但是在Hd1基因编码区、内含子区共检测到43个SNP位点,表明所选择的材料间存在较丰富的遗传变异,具有一定代表性。
CO类基因编码的蛋白质在N末端有含半胱氨酸的锌指结构:C-X2-C-X16-C-X2-C[16],而在C末端有一个CCT域。研究表明,只有发生在这2个区域的变异才会导致CO基因功能的改变[1]。本研究发现,Hd1蛋白在11个谷子品种间存在15个氨基酸替换位点,但是只有2个替换分别发生在锌指结构区和CCT域,1个是第44个氨基酸由酪氨酸突变为半胱氨酸,突变品种为郑州12,1个是第334个氨基酸由精氨酸突变为甘氨酸,突变品种为郑8041(14)。因为这2个突变均为错义突变,可能会导致蛋白质功能的改变,推测郑州12和郑8041(14)对光周期敏感性会减弱。而笔者于2015年、2016年在海南、洛阳、吉林3个不同光周期地区对包含郑州12和郑8041(14)的203个谷子品种抽穗期的调查结果均表明,郑州12具有较弱的光周期敏感性,但郑8041(14)对光周期较敏感(未发表数据),说明Hd1蛋白锌指结构区域的突变对其功能有重要影响,而CCT域包含核定位序列,如果突变发生在该序列,导致不能进行准确的核定位,则会导致基因功能缺失。郑8041(14)并没有对光周期表现弱的敏感性,说明CCT域的突变位点可能没有发生在核定位序列内。
除影响抽穗期外,光周期途径的许多基因还对其他农艺性状产生影响,具有多重效应。如Ghd7基因除影响开花期外,还影响每穗粒数[17]。Hd1基因和Ehd1基因可以减少水稻穗部初级分支数目,使每个穗的小穗数目减少[5]。Hd1基因所在区间除对抽穗期有影响外,还对株高和千粒质量有显著作用[6]。近期的研究还表明,Hd1感光等位基因缺失了光周期敏感性,在自然短日照和长日照下均促进抽穗,并同时降低株高、粒数和产量[7]。目前的研究仅表明,谷子Hd1基因与抽穗日数紧密相关,而在谷子中该基因是否表现多效性尚没有报道。本研究通过Hd1基因与9个农艺性状的初步关联分析发现,该基因除了与抽穗日数关联外,与千粒质量也存在明显关联,同时还与株高和穗粗相关,表现多效性,与在水稻中的研究结果[6-7]存在一定吻合,这表明不同植物CO基因在功能上具有相似性。由于本研究所用的谷子材料有限,因此要想更充分证明Hd1基因的功能,在今后的进一步研究中需要扩大品种数量,获得更多的表型变异类型,充分发掘Hd1-like基因存在的突变,以使关联结果更可靠。
通过调查11个谷子品种抽穗日数、株高、穗长、穗粗、穗质量、码粒数、穗码数、穗粒质量、千粒质量9个农艺性状,对除抽穗日数之外的8个性状进行品种间差异显著性检验表明,所有8个性状在11个谷子品种间均存在显著差异,株高、穗长、穗质量等多数性状存在丰富的变异类型。对所有9个性状进行相关性分析表明,株高、抽穗日数与多数穗部性状存在显著或极显著正相关,而穗部各性状是构成谷子产量的重要因子。因此,株高和抽穗日数对谷子产量潜能具有重要影响。首次克隆了11个谷子品种的Hd1-like基因序列,序列比对发现,该基因编码区、内含子区存在43个变异位点,且都为SNP,未发现插入/缺失突变;推定的Hd1蛋白序列在11个谷子品种间检测到15个氨基酸替换位点,其中2个替换发生在保守的锌指区域和CCT结构域,而锌指结构区的突变可能导致了郑州12对光周期敏感性减弱。初步进行了突变位点与农艺性状的关联分析,发现谷子Hd1-like基因为一个多效基因,除了与抽穗日数、千粒质量存在关联,还与株高、穗粗存在关联。
