冬小麦初生根与次生根形态、生理性状差异分析
2018-08-02臧贺藏王言景卢红芳贺德先
臧贺藏,王言景,张 均,李 丰,卢红芳,贺德先**
(1.河南省农业科学院 农业经济与信息研究所,河南 郑州 450002;2.河南农业大学 农学院,河南 郑州 450002; 3.郑州师范学院 初等教育学院,河南 郑州 450044)
在小麦生长发育和产量形成过程中,初生根和次生根相互补充、相互促进[1]。前者分化形成时间早、数量少,对植株根系、生长发育及稳产性等起重要作用[2-3];而后者发生时间长、数量多,对巩固和协调初生根,实现高产具有决定作用[4]。根系的生长具有可塑性,其功能的发挥与根系形态和生理特性密切相关,因而,协调初生根与次生根及其与产量的关系一直是作物学科极为关注的热点。
初生根、次生根的发育受其所处的环境和相关栽培措施影响,最主要的是由遗传因素决定,且初生根的发育较次生根稳定[5-7]。有研究表明,不同基因型小麦的初生根和次生根在根数、根体积、根长、根呼吸及根干质量方面表现不同,前期初生根较好的发育和较高的根系吸水功能有助于促进次生根较好建成[8]。也有研究表明,初生根受土壤水分影响较小,而次生根对土壤水分极为敏感[9]。还有研究指出,初生根、次生根的建成和功能不同,初生根下扎深,可以从深层土壤中吸收水分,对小麦的抗旱性起着至关重要的作用;而次生根分布较浅,主要吸收利用浅层土壤水分[10-12]。小麦初生根与次生根的形态、生理特征是根系质量的重要体现,与植株的生长发育、养分吸收、产量形成等密切相关。目前,关于小麦[13-15]、玉米[16-21]、水稻[22-26]、高粱和小米[27]、甘薯[28]等作物的根系形态研究较为系统,但文献资料查阅结果表明,迄今尚未见在高产条件下对小麦全生育期内初生根和次生根形态数量和生理性状差异的系统报道,而这样的研究对河南省小麦高产、高效生产具有重要的指导性。适当调控初生根与次生根的数量,发挥初生根与次生根功能,维持生育后期初生根与次生根的生理活性,有利于综合提高产量。鉴于此,以河南省小麦为材料,采用分根移位的方法,重点探讨不同生育时期初生根和次生根的形态和生理特征,并揭示二者之间的差异,旨在为有针对性地采取措施调控初生根和次生根的生长发育,充分发挥2类根系的生理功能,提高根群质量和根系生理势,最终为高产、高效、优质栽培提供科学依据。
1 材料和方法
1.1 试验材料及试验地概况
供试材料为河南省小麦品种:郑麦9023(弱春性,强筋,早熟)、郑麦004(半冬性,弱筋,抗寒)、周麦16号(半冬性,中筋,抗寒)、洛旱2号(半冬性,中筋,抗旱)、兰考矮早8(弱春性,大穗大粒,综合抗性好)、豫麦18号(弱春性,中筋,抗倒)。
试验在郑州河南农业大学科教园区进行,该区属温带半湿润季风气候,前茬田菁掩底。供试土壤为砂质壤土,0~40 cm耕层土壤养分含量为:全氮0.99 g/kg、碱解氮57.9 mg/kg、有效磷44.4 mg/kg、有效钾204.8 mg/kg、有机质17.8 g/kg、pH值7.9。
1.2 试验设计
试验采用盆栽分根移位法进行研究,盆栽用塑料盆直径为0.23 m,高为0.25 cm;将耕层土壤过筛,每盆装土10 kg,基施尿素1.2 g、过磷酸钙2.4 g和氯化钾0.8 g;在拔节期每盆追施尿素1.2 g。10月26日大田播种,11月4日分根移栽,每盆定植10株,重复4次。
1.3 试验方法
1.3.1 分根移位法 分根移位法是在保证小麦幼苗正常生长条件下,将初生根、次生根分开并引到不同生长环境中培养的一种研究方法。该法首先在田间预留地上播种,播种稍微深些,便于区分出初生根和次生根。盆中土壤的填装过程如下:装土前,使用塑料薄膜将塑料桶中间隔开,并在塑料薄膜约距土壤上表面6 cm处,打出直径为3 mm的小孔,盆内暂装土到小孔下缘处并压实。幼苗移栽:用铁锹挖出带土幼苗,选择带有地中茎的幼苗进行移栽。将幼芽和地中茎穿过小孔,初生根分隔于小孔的另一侧,扶正幼苗,然后在两侧覆土,覆土厚4~5 cm,灌水。
1.3.2 根系样品采集 分别于11月20日(冬前分蘖期)、12月15日(越冬前)、2月5日(返青期)、3月15日(拔节期)、4月15日(挑旗期)、5月5日(籽粒形成期)、5月15日(灌浆中期)、5月28日(成熟期)进行根系取样,然后采用浸泡—筛选—复拣的方法进行冲洗。首先,选择在池子里充分浸泡的盆栽植株,将土壤泡成泥状,冲洗时根、土容易分离;然后,反复冲洗,清除掉附着于根上的泥土、半腐解杂物等;最后,再取1个装有水的大塑料盆,将根系置于塑料盆中,用筛网捞去或人工拣去半腐解杂物等。
1.3.3 根系形态及生理指标的测定 根数采用目测法,直接计数。单株根干质量:将洗净的根在80 ℃条件下烘至恒质量,冷却后用千分之一天平称质量。根系活力采用改良TTC法测定。可溶性糖含量采用蒽酮法测定。全氮含量采用全自动凯氏定氮仪测定。
1.3.4 产量及其构成因素 成熟时,每个小麦品种取4盆进行考种、计产。
1.4 统计分析方法
试验数据使用Excel 2010和SPSS 19.0进行统计分析。
2 结果与分析
2.1 冬小麦根数与产量及其构成因素的关系
初生根与次生根生长状况直接影响冬小麦地上部形态建成和籽粒产量的高低,根数与产量的关系较为密切。由表1可知,对于单株初生根数来说,郑麦9023、郑麦004与周麦16、洛旱2号、兰考矮早8之间的差异显著;对于单株次生根数来说,除郑麦9023与周麦16之间的差异显著外,其他品种间的差异均未达到显著水平。从产量及其构成因素来看,不同小麦品种单株产量介于2.28~2.72 g,其中郑麦9023产量最高,兰考矮早8产量最低;不同小麦品种千粒质量介于40.0~45.6 g,郑麦9023千粒质量最高,洛旱2号最低;不同小麦品种穗粒数表现为:郑麦004>郑麦9023>洛旱2号>周麦16>兰考矮早8>豫麦18;不同小麦品种单株穗数表现为:郑麦004>郑麦9023>豫麦18>周麦16>洛旱2号>兰考矮早8。以上结果说明单株初生根数越多,千粒质量和产量越高,单株初生根数对产量的贡献高于单株次生根数。
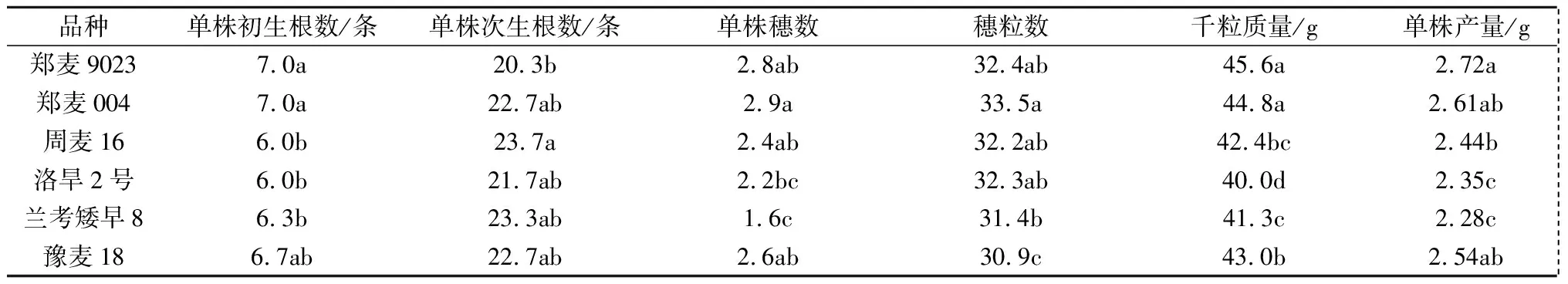
表1 冬小麦根数、产量及其构成因素
注:同列数据后不同小写字母表示不同品种之间差异显著(P<0.05)。
由表2可以看出,单株初生根数与千粒质量、单株产量均呈显著正相关,相关系数分别为0.888、0.840,与单株穗数和穗粒数无显著相关;单株次生根数与千粒质量、单株产量、单株穗数和穗粒数均无显著相关性。说明单株初生根数对产量的作用明显大于单株次生根数,同时单株初生根数对产量的影响主要通过调节千粒质量而起作用。
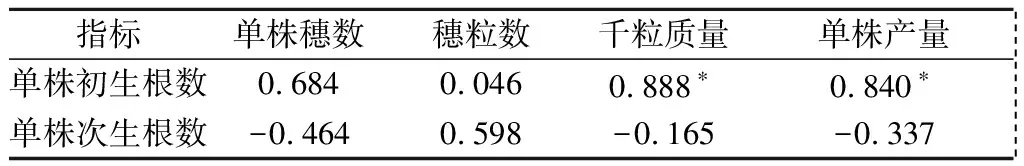
表2 冬小麦根数与产量及其构成因素的相关性
注:*表示在0.05水平上显著相关。
2.2 不同生育时期冬小麦初生根与次生根形态性状的差异
2.2.1 单株根数 单株初生根数受环境影响较小,能够吸收利用土壤深层水肥,对小麦高产稳产具有重要意义。单株次生根数既受基因型的影响,又对生态条件反应敏感,环境适宜时发生数量多,不适宜时则较少,其数量多少在一定程度上直接反映了小麦植株个体的健壮程度和丰产性。由表3可以看出,随着生育进程的推进,冬小麦单株初生根数逐渐降低,介于6.5~7.6条;单株次生根数呈先升高后降低的变化趋势,挑旗期达到高峰,为46.2条。经分析发现,各生育时期单株初生根数与单株次生根数的差异均达显著水平。其中,拔节以前以初生根生长为主,单株初生根数占单株总根数的比例较高,冬前分蘖期为80.9%;随着生育进程的推进,单株次生根数迅速增多,单株初生根数所占比例逐渐降低,至成熟期仅为23.7%。因此,在高产优质栽培实践中,小麦需要根群数量适宜而生理功能强大的初生根系和次生根系。

表3 不同生育时期冬小麦单株根数的差异 条
注:同行数据后不同小写字母表示不同时期之间差异显著(P<0.05),同列数据后不同大写字母表示不同根系类型之间差异显著(P<0.05),下同。
2.2.2 单株根干质量 由表4可知,冬小麦单株初生根干质量随生育进程的递进呈先升高后降低的变化趋势:冬前分蘖期至返青期为0.012~0.061 g,拔节期至灌浆中期为0.121~0.334 g,灌浆中期以后部分初生根衰亡,干质量出现负增长,至成熟期为0.240 g。由此可见,不同品种单株初生根干质量在灌浆中期达到最大值,成熟前仍维持较高水平,充分说明初生根在全生育期内自始至终都起作用,并部分解释了初生根与稳产性之间的密切关系。单株次生根干质量随生育进程的递进呈先升高后降低的变化趋势:冬前分蘖期至返青期,单株次生根干质量为0.002~0.027 g;拔节期至灌浆中期,单株次生根干质量为0.156~0.789 g;成熟期,单株次生根干质量为0.702 g。
另外分析发现,单株初生根干质量与单株次生根干质量之间的差异在各生育时期均达到显著水平。生育前期,单株初生根干质量占单株根干质量的比例较高;随着生育进程的推进,单株初生根干质量所占比例逐渐降低,而单株次生根干质量所占比例升高(表4)。说明在生育前期保证初生根较多生长条件下,促进次生根较早出现对总根量的形成有益。
2.3 不同生育时期冬小麦初生根与次生根生理性状的差异
2.3.1 根系活力 由表5可知,随生育进程的推进,冬小麦初生根活力逐渐增强,至返青期达最大值,为254.745 μg/(g·h);返青期以后,初生根活力逐渐减弱,至灌浆中期出现次高峰,初生根活力为84.692 μg/(g·h);之后,到成熟期初生根活力降至最低。冬小麦次生根活力随生育进程的推进逐渐升高,到拔节期达最大值,为329.092 μg/(g·h);之后呈下降趋势,至灌浆中期出现次高峰,成熟期降至最低。另外,经分析发现,在各个生育时期,小麦初生根活力均显著低于次生根。在生产实践中,应注重提高初生根生理活性,维持初生根、次生根活力稳定至生育后期,进而提高产量。

表5 不同生育时期冬小麦根系活力的差异 μg/(g·h)
2.3.2 全氮含量 由表6可以看出,随生育进程的推进,小麦初生根全氮含量呈降低趋势,在返青期最高,为28.970 mg/g,拔节期至灌浆中期为17.048~23.537 mg/g,成熟期为16.352 mg/g。次生根全氮含量与初生根全氮含量的变化趋势相同,返青期为34.375 mg/g,拔节期至灌浆中期为11.885~25.532 mg/g,成熟期为10.096 mg/g。返青期至拔节期初生根全氮含量显著低于次生根;挑旗期至成熟期初生根全氮含量显著高于次生根。
2.3.3 可溶性糖含量 冬小麦初生根可溶性糖含量随生育进程的推进呈现降低趋势(表7),至成熟期降为7.039 mg/g。次生根可溶性糖含量的变化趋势与初生根一致,至成熟期降为3.676 mg/g。冬小麦初生根与次生根可溶性糖含量的差异在各个生育时期均达显著水平。其中,返青期至挑旗期初生根可溶性糖含量显著低于次生根,而籽粒形成期至成熟期初生根可溶性糖含量则显著高于次生根。

表6 不同生育时期冬小麦根中全氮含量的差异 mg/g

表7 不同生育时期小麦根中可溶性糖含量的差异 mg/g
3 结论与讨论
小麦籽粒产量的高低取决于根系的生长发育状况,发达的根系和较高的根系生理活性是小麦高产稳产的基础[29-31]。有研究认为,高产小麦应具有根系发达、发根力强、根数多、扎根深,根系活力较强、吸收养分和水分的能力较强、后期衰老较晚且缓慢等特点[32-34]。也有研究认为,小麦深层初生根、次生根生长易受滴灌量少、湿润土层浅的影响,且根系分布浅,初生根提前衰老,导致千粒质量降低而减产[35]。而另有研究认为,初生根数量和质量对小麦籽粒产量的形成具有贡献,初生根对产量的相对作用明显高于次生根[36-37]。本研究也持有相同看法,认为单株初生根数与千粒质量、产量呈显著正相关,说明单株初生根数对产量的贡献显著高于单株次生根数。然而,利用初生根数预测籽粒产量具有一定局限性,这是因为忽视了强大的次生根系统及其与环境的互作。本研究发现,小麦初生根与次生根形态数量性状的差异均达显著水平:冬前分蘖期至返青期,单株初生根数和单株初生根干质量显著高于单株次生根;返青期之后,单株次生根数和单株次生根干质量显著高于单株初生根。生产实践中,在保证生育前期初生根系较快、较多生长的前提下,促进次生根早发快长对实现小麦高产、优质、高效栽培具有重要作用。
根系是吸收水分和养分的主要器官,其生理活性的强弱与作物地上部的生长发育及产量形成等密切相关,但不同品种对根系生理活性的反应存在明显差异[38-39]。小麦根系生理活性在全生育期内的变化幅度很大,尤其是生育后期初生根和次生根的生理活性均不强盛、功能维持时间短,是造成产量波动的重要原因,因而如何提高生育后期初生根和次生根的生理活性,是实现高产、高效、优质栽培的关键所在。有研究表明,根系中的老根所占比例随生育进程的推进逐渐增大,尤其生育中后期根系活力的衰退速率逐渐加快[33]。也有研究表明,抗旱性强的小麦品种深层根系具有较高的活力和较强的吸水能力,进而利于增加粒质量[40]。本研究表明,不同生育时期冬小麦初生根与次生根生理性状的差异均达显著水平。从各生育阶段根系活力来看,初生根显著高于次生根。生育前中期,全氮含量和可溶性糖含量均表现为初生根显著低于次生根;生育后期,表现为初生根显著高于次生根,这说明生育后期次生根物质向外转运较多,与马冬云等[41]的研究结果一致。比较而言,较强的初生根生理活性有助于促进次生根的较好建成。通过栽培措施调控初生根与次生根,选用根系活力高的小麦品种将更有利于提高产量。
