尖镰孢胁迫下黄瓜茎部蛋白质差异表达分析
2018-08-01祝海娟杜洪锐陈瑞雪党悦嘉周莹莹张艳菊
杨 森, 祝海娟, 杜洪锐, 陈瑞雪, 党悦嘉, 周莹莹, 张艳菊*
(1. 东北农业大学农学院, 哈尔滨 150030,2. 哈尔滨市阿城区种子服务中心, 哈尔滨 150030)
植物在逆境胁迫环境下,一些基因被诱导表达,实现植物在蛋白组学水平的适应性调整,构建了植物整个复杂且高效的响应系统。大量研究表明,病程相关蛋白(pathogenesis related protein)是主要的宿主功能蛋白,这些重要的功能蛋白在不同类型互作系统中得到了充分的验证,明确了各种类型植物防御机制相关功能性蛋白的作用[1-3]。利用蛋白质组学技术构建植物宿主-病原菌互作体系已经成为当前研究的趋势和热点之一。
由无性型真菌镰孢属尖镰孢黄瓜专化型Fusariumoxysporumf.sp.cucumerinum侵染引起的黄瓜枯萎病是黄瓜生产中的重要土传病害,由于保护地的特殊环境条件和长期连作,土壤中病原菌的积累量越来越多,导致黄瓜枯萎病等土传病害发生逐年加重。F.oxysporum通常定殖于植物的维管束,对输导组织产生危害,同时产生相应的毒素,致使茎基部维管束变褐,严重地影响了水、矿物质及营养的运输及黄瓜的生长[4]。植物茎部除了运输水、矿物盐、营养和代谢物外,也参与长距离信号运输,作为对病原菌、共生体和环境压力入侵的一种反应场所[5]。当病原菌侵染植物时,会有病程相关蛋白及其产物等一系列抗性相关物质产生,其中包括代谢酶、与压力有关的蛋白质等。
本研究采用双向电泳技术分析在两种毒力不同的尖镰孢胁迫下黄瓜抗感枯萎病品系茎中蛋白质的差异表达,用质谱分析方法鉴定与抗病相关的蛋白质,从蛋白水平初步探索抗枯萎病机制,从而为维管束病害中寄主与病原的互作提供理论依据。
1 材料与方法
1.1 试验材料
黄瓜枯萎病高抗品系‘D0327’和感病品系‘649’,由东北农业大学黄瓜课题组提供。尖镰孢黄瓜专化型Fusariumoxysporumf.sp.cucumerinum强毒力菌株C9和弱毒力菌株S1,由东北农业大学植物病理研究室提供[6]。
1.2 试验方法
1.2.1 接种
将培养在PSA平板上的供试菌种移入PL培养液中,置于25~28℃,转速110~120 r/min的恒温摇床中振荡培养7~10 d。将菌液用4层纱布滤掉菌丝体,滤液在转速4 000 r/min下离心10 min后移去上清液,加入无菌蒸馏水配制成1.0×107个/mL的孢子悬浮液。于黄瓜幼苗两片真叶期,采用灌根法将2个毒力不同的尖镰孢菌株分别接种抗感黄瓜品系[7]。接种后待感病品系‘649’茎部出现变黄症状后(7 d)进行取材,将整株茎剪成5 cm茎段用锡纸包好,迅速用液氮处理后置于-80℃冰箱中保存备用。
1.2.2 黄瓜茎部蛋白质的提取
利用TCA-丙酮法[8],提取黄瓜茎组织蛋白质。利用考马斯亮蓝法[9]测定蛋白质浓度并进行定量。称取1 g黄瓜茎放入预冷的研钵中,加入液氮研磨至粉末状;将粉末转移至2 mL EP管中,加入5倍体积预冷的10%TCA-丙酮溶液,漩涡振荡至其混合充分,放于-20℃冰箱沉淀过夜; 4℃,13 000 r/min离心30 min,移去上清液;向沉淀中加入5倍体积预冷的丙酮溶液(含0.07%β-巯基乙醇,-20℃预冷),充分混合,置于-20℃冰箱中静置1 h(期间漩涡振荡几次); 4℃,13 000 r/min离心30 min,移去上清液(重复上述步骤6~7次,洗至上清呈无色);沉淀经真空干燥制成蛋白质干粉,-80℃保存备用。
1.2.3 蛋白质双向电泳
第一向电泳选择24 cm,pH 4~7的IPG胶条进行等电聚焦,第二向电泳采用12.5%的SDS-PAGE分离胶,双向电泳结束后进行考马斯亮蓝G-250染色,每个处理进行3次重复。获得的2-DE凝胶图像通过Imagemaster 2D Platinum 7.0软件进行图像分析。与参考胶比较,特异表达、表达量上调或下调2倍以上被认为是差异表达的蛋白质点。
1.2.4 质谱鉴定和蛋白鉴定
委托哈尔滨赛信生物科技有限公司进行MALDI-TOF-TOF/MS质谱分析。质谱分析结果利用软件Mascot 2.3.02进行分析,经过NCBI(nr)数据库和黄瓜基因组数据库(http:∥www.icugi.org/cgi-bin/ICuGI/genome/index.cgi?organism=cucumber)分析,获得相关蛋白质点的鉴定结果。
1.2.5 差异蛋白基因表达验证
在黄瓜基因组数据库中搜索编码蛋白的基因,利用在线工具GenScript Real-time PCR (TaqMan) Primer Design进行设计(https:∥www.genscript.com/ssl-bin/app/primer)。引物序列发送至上海生工生物工程技术服务有限公司进行合成,引物序列见表1。
表1qRT-PCR引物序列
Table1PrimersequencesforqRT-PCR

基因编号Gene no.引物序列(5'-3')Sequence of primerCsEF1αF:CGCTCTTCTTGCTTTCACCCTTR:TACCTTGCCTTGGAGTATTTGGCsa5M611600.1F:GCAACACTTGGCCGTATCATR:AATTTGCTGTTCCGTTGCCTCsa6M484600.2F:CACCATCGGAGCTGAGAGATR:CTCATCCTGTCTGCAATGCCCsa4M063440.1F:TACAGCGTCCAACGTGAATGR:CCCTCCACAAACTTCCTTGCCsa6M450370.1F:TGGTGCAAATGCCATACTCGR:TGCAAGCTTGTTTCCTGCATCsa4M598000.1F:TGGAACAAACGAGGAGGTGAR:CCAGTAAACGCACCACCTTTCsa3M116710.1F:TGGACTCTGATGTGGGAACCR:GTAAACGGGACCTTCTGGGACsa017651F:GGGTGATCTATATGGAAAGCTGGAR:TTCATGTAGGTCGACACCGTCsa6M522690.1F:CCTCTGCAATGGCTCTTTCCR:CTGACTTGCTGGCAGTCTTC
采用TRIzol法提取黄瓜叶片总RNA[10]。根据Toyobo反转录试剂盒ReverTra Ace qPCR RT-Kit说明书反转录成cDNA。依照SYBR®Green Realtime PCR Master Mix说明书进行。参照基因为CsEF1α(GenBank Accession Number: XM_004138916)[11]。反应体系SYBR Green PCR Master Mix, 10 μL; 上游引物0.5 μL; 下游引物0.5 μL; cDNA 2 μL; ddH2O 7 μL。反应条件:95℃ 3 min;95℃ 10 s,58℃ 20 s,72℃ 30 s,40次循环;55℃ 10 s,81次循环;4℃,10 min。采用2-ΔΔCT相对定量分析方法计算出基因的相对表达量[12],并用DPS 7.05数据处理系统软件进行方差及显著性分析。
2 结果与分析
2.1 尖镰孢胁迫下不同抗性黄瓜品系茎组织蛋白双向电泳图谱分析
2.1.1 C9胁迫下黄瓜抗、感品系蛋白双向电泳图谱
利用Imagemaster 2D Platinum 7.0图像软件分析,并对分析结果进行筛选,最终确定C9胁迫下感病品系中共有6个符合2倍以上并且有99%统计学显著性差异的蛋白质点,抗病品系中共有3个差异蛋白点(见图1)。

图1 C9胁迫下黄瓜抗、感品系茎部蛋白的双向电泳图Fig.1 The 2-DE map of differential protein spots in susceptible and resistant cucumber lines under the stress of C9
2.1.2 S1胁迫下黄瓜抗、感品系蛋白双向电泳图谱
S1胁迫下感病品系中共有4个符合2倍以上并且有99%统计学显著性差异的蛋白质点,抗病品种中共有5个差异蛋白点(见图2)。从双向电泳图谱可以看出,无论感病还是抗病品系,S1胁迫下出现的差异蛋白点大都集中在对照。

图2 S1胁迫下黄瓜抗、感品系茎部蛋白的双向电泳图Fig.2 The 2-DE map of differential protein spots in susceptible and resistant cucumber lines under the stress of S1

点号Spot no.基因编号Gene no.分子量/DaMW等电点pI序列覆盖率/%Sequence coverage得分Score蛋白名称Protein name表达模式Expression pattern1Csa5M611600.159 885.25.9036470Enolase上调2Csa6M484600.241 911.05.31721 100Actin上调3Csa4M063440.128 292.37.6144557Oxygen-evolving enhancer protein下调4Csa6M450370.147 942.55.4861752Enolase isoform X1上调5Csa4M598000.127 502.35.6165784Triosephosphate isomerase上调6Csa3M116710.116 450.56.3073657Nucleoside diphosphate kinase上调7Csa01765118 010.95.1544233Csf-2 protein下调8Csa6M522690.128 239.15.1430258Chlorophyll a-b binding protein of LHCII type I上调
2.2 差异蛋白质点的质谱鉴定
通过质谱技术检测,获得质谱图谱,8个PMF出峰情况良好,根据NCBI黄瓜数据库检索得到8个阳性结果。通过对两种尖镰孢处理后的抗感黄瓜品系进行分析,最终成功鉴定出8个差异蛋白点,其中6个差异蛋白上调表达,2个下调表达,如表2。差异蛋白点的质谱结果如下。
强毒力菌株C9胁迫下,感病品系中鉴定出3个差异蛋白点,分别是1、2和3号,抗病品系中差异蛋白为4、5及6号。弱毒力菌株S1胁迫下,感病品系中差异蛋白为7号,抗病品系中差异蛋白为8号。
1和4号为烯醇化酶及其亚基。在糖酵解中烯醇化酶及其亚基主要参与2-磷酸-D-甘油酸与磷酸烯醇式丙酮酸之间进行相互转化。
2号蛋白点为肌动蛋白。肌动蛋白是构成真核生物细胞骨架的重要成分,它参与细胞运动,以及多种细胞内过程,对细胞生长、分裂、分化、细胞内囊泡运输、细胞壁的生物合成、共生现象、胞吞胞吐作用以及膜的循环利用等方面具有较好的控制作用[13]。
3号和8号蛋白点分别为放氧增强蛋白OEE (oxygen-evolving enhancer protein)和捕光复合体I型叶绿素a/b 结合蛋白(LHCII type I chlorophyll a/b-binding protein)。这2个蛋白均与植物光合作用有关,8号蛋白参与光合作用原初反应的蛋白质,主要功能进行捕获叶绿素,相关研究指出叶绿素的含量与植物的抗病性呈正相关,叶绿素含量升高,植株的抗病性增强。
5号蛋白点为磷酸丙糖异构酶(TPI)。磷酸丙糖异构酶在生物体内的功能主要是参与糖酵解,是糖酵解途径中的主要蛋白酶。TPI催化磷酸丙糖异构体在二羟丙酮磷酸和D-甘油醛-3-磷酸之间的可逆转化,是糖酵解中不可或缺的酶。
6号蛋白点为核苷酸二磷酸激酶(NDPK/NDK)。在生物体内核苷酸二磷酸激酶催化磷酸基团在ATP和NDP两者之间进行可逆的转移,用以维持ATP和NTP两者浓度在细胞内的稳定。
7号蛋白点为Csf-2蛋白。该蛋白是一种分泌蛋白,其主要功能与植物防御反应相关,参与应答生物刺激,但具体作用机理尚未清楚。
2.3 差异蛋白基因表达验证
通过qRT-PCR对差异蛋白基因的表达进行了验证,结果如图3所示。8个差异蛋白基因的表达模式均与其蛋白表达相一致,说明差异蛋白是由基因差异表达引起的。
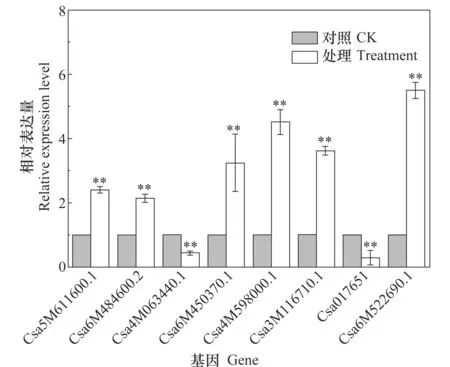
图3 差异蛋白基因相对表达量 Table 3 Relative expression of differential protein genes
3 讨论
蛋白质是生命最直接的体现,在生理及病理条件的改变下,生物体的蛋白会产生结构及功能的变化[14]。植物在被病原物侵染刺激后,其本身会产生一系列的防御反应,尤其是一些病程相关蛋白的表达会有一定的变化[15-16]。本研究采用双向电泳技术分析在两种毒力不同的尖镰孢胁迫下黄瓜抗、感枯萎病品系茎中蛋白质的差异表达,共鉴定出8个差异蛋白。根据其功能不同,将8个蛋白分为能量代谢相关蛋白、光合作用相关蛋白和胁迫反应相关蛋白。
能量代谢相关蛋白为1、4、5、6号,1、4号蛋白为烯醇化酶及其亚基,5号蛋白为磷酸丙糖异构酶(TPI),6号蛋白为核苷酸二磷酸激酶(NDPK/NDK),这4种参与能量生成的蛋白质在本研究中均出现上调表达,且3种在抗病品系出现,说明在同一菌株胁迫下抗感品系表现出一定的差异性,对外界不良环境表现出一定的抗性。这4种与能量相关的蛋白均在高毒力菌株C9的胁迫下差异表达,说明毒力高的菌种可更好地激活植物组织相关蛋白的表达,进而提高抗病性。
光合作用相关蛋白为3、8号,3号蛋白为放氧增强蛋白OEE (oxygen-evolving enhancer protein),8号蛋白为捕光复合体I型叶绿素a/b 结合蛋白(LHCII type I chlorophyll a/b-binding protein)。叶绿素含量与植物抗病性正相关[17],8号是参与光合作用原初反应的蛋白质,与植物叶绿素含量相关,很可能参与黄瓜抗枯萎病的反应。S1菌株胁迫下,8号蛋白在抗病品系中上调表达,而C9菌株胁迫下,3号蛋白在感病品系中下调表达,说明在毒力不同的菌株诱导下蛋白表达存在差异。
胁迫反应相关蛋白为2号,肌动蛋白。该蛋白在高毒力菌株侵染感病品系中上调表达。肌动蛋白普遍存在真核生物中,在细胞外表组织形态的维持、正常生长、细胞运动等中具有重要意义[9,18]。
通过本试验可以看出在病原菌胁迫下,为适应外界环境的变化,植物体内迅速做出反应,尤其是在毒力强的菌株胁迫下表现得更为明显。通过分析上述试验结果,说明参与物质和能量代谢的蛋白与黄瓜抗枯萎病相关。本研究初步探讨了黄瓜枯萎病菌诱导黄瓜的抗性应答机理,为进一步筛选抗性相关特异蛋白质作为标记性状选育新的抗性黄瓜品种奠定理论基础。
