新疆地方绵羊ND2基因多态性及系统发育分析
2018-08-01王世锋米耶斯尔詹建立张秀英王玉涛
王世锋, 米耶斯尔, 詹建立, 张秀英, 王玉涛
(喀什大学生命与地理科学学院叶尔羌绿洲生态与生物资源研究重点实验室,新疆喀什 844000)
绵羊与人类生活密切相关,为人类提供了肉、脂、奶、皮、毛、绒、骨和角等多种多样的畜产品[1-3]。关于绵羊的起源仍存在争议,线粒体DNA(mitochondrial DNA,mtDNA)序列分析已经确定了一种多母系血统的普遍现象,即A、B、C、D、E进化世系,这些世系有特定的地理范围,暗示多重母系起源,羊驯化事件可能是独立的[4]。目前,研究发现仅欧洲摩弗伦羊可能是家绵羊的母系(世系B)野生祖先之一。推测亚洲摩弗伦羊与欧洲摩弗伦羊与家绵羊共享最近的母系祖先[5],未发现其他野生羊对家绵羊有遗传贡献的分子证据。
线粒体是真核细胞的动力车间,mtDNA作为独立与核DNA的半自主性基因组,是核外遗传物质,动物体线粒体基因组编码的37个基因,包括13条多肽,22个tRNA基因和2个rRNA基因[6],具有分子量小、进化速度快、多挎贝、碱基替换率低、遗传自主及母系遗传等特征。
ND2基因是mtDNA的1个蛋白编码基因,而且是NADH脱氢酶的1个亚基,而NADH脱氢酶是呼吸链复合体1的主要组成,在呼吸链中直接参与氢与电子传递并通过氧化磷酸化产生ATP,进而参与能量代谢作用。NAD参与生物体内的重要反应,NAD可在糖酵解途径与三羧酸循环途径得到电子形成NADH,NADH又在氧化磷酸化途径中将电子通过呼吸链最终传递给氧,促成ATP的形成[7]。在体内大多数代谢脱下的氢,是由NADH呼吸链传递给氧,糖类碳水化合物、脂质和蛋白质的分解代谢,脱氢后的氧化反应一般是由NADH呼吸链传递完成的。
鲁卫卫等报道鸡线粒体ND2基因的异质性,并发现异质性变异对生长性状、屠体指标、血清生化等指标均有显著效应,显示相关变异的潜在重要性[7]。鲍海港等在对藏鸡的研究中发现,线粒体ND1和ND2基因可被选为藏鸡低氧遗传适应的候选基因,因为低氧会抑制与线粒体呼吸功能密切相关的酶的活性,也会使线粒体产生更多的活性氧。活性氧会破坏膜脂,降低线粒体呼吸酶的活性,从而损害线粒体的呼吸功能。研究发现,3种鸡都有但频率差异较大的2个错义突变都是ND2基因上的单碱基突变,推测藏鸡与低地鸡有可能在NADH脱氢酶的功能上存在差异[8]。而有关高山放牧品种的塔什库尔干羊(海拔3 000~5 000 m)、和田羊(分为山区型和农区型,海拔1 440~3 500 m)和多浪羊(海拔1 200 m)线粒体呼吸功能和低氧适应研究较少。
本研究通过测定绵羊线粒体ND2基因序列,从而探讨新疆南疆地区绵羊的遗传多样性、线粒体呼吸功能及低氧适应的分子机制,从而了解并摸清其亲缘关系及低氧适应性,为能够综合开发和合理利用绵羊资源提供一定的科学依据。
1 材料与方法
1.1 试验动物
于2014年3月至2015年8月先后在麦盖提县种羊场采集33只多浪羊、塔什库尔干县麻扎种羊场采集41只塔什库尔干羊、和田市屠宰场采集17只和田羊共计91只羊的静脉血液,用柠檬酸葡萄糖(ACD)抗凝,冷冻后置于冰袋中,于实验室-20 ℃保存备用。
1.2 主要试剂
粪便基因组DNA快速提取试剂盒、全血基因组DNA快速提取试剂盒,均购自北京三博远志生物技术有限责任公司;TaqDNA聚合酶、dNTP,均为杭州博日科技有限公司产品;琼脂糖为BIOWEST公司产品。
1.3 总DNA提取
绵羊总基因组DNA的提取是利用全血基因组DNA快速提取试剂盒(北京三博远志生物技术有限责任公司)。
1.4 PCR扩增及测序
根据绵羊mtDNA序列(GenBank登录号为AF010406),采用Primer 5.0软件设计引物,扩增绵羊ND2基因,上游引物:5′-AGCACCCACTGATTGCTCAT-3′,下游引物:5′-TTCGTTTTGTGGTTGGGAAT-3′。委托北京三博远志生物技术有限责任公司合成,期望扩增长度约901 bp。
PCR反应体系总体积为50 μL:绵羊总基因组4 μL,10×Buffer(含Mg2+)5 μL,上下游引物各2 μL,2.5 mmol/L dNTPs 4 μL,TaqDNA聚合酶1 μL,去离子水29 μL。
PCR反应条件:94 ℃预变性4 min;94 ℃变性30 s,53 ℃退火30 s,72 ℃延伸1 min,进行35个循环;最后72 ℃再延伸10 min,4 ℃保存。
PCR扩增产物直接送北京三博远志生物公司测序。
1.5 数据分析
应用Clustal X 1.83软件进行序列比对并手工校对。DnaSP 5.1软件计算单倍型多样度、核苷酸多样度、多态位点、核苷酸总变异位点、简约信息位点和点突变位点等。MEGA 5.1 软件分析变异位点,计算碱基组成、转换/颠换比(Ts/Tv)、核苷酸差异和序列差异,采用邻接法(Neighbour-Joining,简称NJ)Kimura双参数模型构建系统发生树,对拓扑图进行重复抽样1 000次的自展检验(Bootstrap)以确定各分支的置信度。
2 结果与分析
2.1 ND2基因PCR扩增结果
ND2基因扩增产物经1.5%琼脂糖凝胶电泳20 min,结果见图1。

电泳条带明亮,扩增效果较好,扩增条带大小为900 bp左右,与预期扩增的901 bp一致。可以直接测序。
2.2 绵羊mtDNA-ND2全序列碱基分析
测序结果表明,所测出的绵羊mtDNA-ND2基因A、T、C、G含量在绵羊中分布较一致,其中A占36.8%,T占 27.4%,C占27.1%,G占8.7%。绵羊的A+T含量为 64.2%,C+G含量为35.8%,A+T含量高于C+G含量;表明绵羊mtDNA-ND2基因含有大量的碱基A和T,存在一定的碱基偏倚性。碱基组成表明G相对缺乏,其中密码子第3位上G的含量最低,仅为2.6%;第3位的A含量较高,为49.8%。密码子的使用存在一定差异,在密码子的第1位上A高达43.4%;密码子第2位上T的含量高达45.0%,碱基G的含量为9.1%。
2.3 绵羊mtDNA-ND2序列的多样性分析
利用DnaSP 5.1软件进行单倍型统计分析,本试验分析91个绵羊mtDNA-ND2基因序列,发现了22个多态位点,其中有9个单一多态位点,简单信息位点(2个碱基)有13个(图2)。其中438、504、837、873位点核酸变异只出现在塔什库尔干羊个体中。核酸变异中转换远大于颠换,转换/颠换比为78.38。

2.4 ND2氨基酸分析
22个多态位点中的2个位点即299、883位点是非同义突变,分别导致M变为T和R变为G,其余20个位点为同义突变。应用SWISS-MODEL(httP://swissmodel.expasy.org/interactive# structure)软件进行ND2结构在线预测,多肽空间结构见图3。
单倍型H1序列未发生突变,H8序列在299位点和883位点均发生突变。由图3可知,突变后空间结构差异不大。但是QMEAN分别为-4.35和-4.38,Cβ分别为-1.72和 -1.82,All Atom分别为-1.27和-13.2,Torsion分别为 -4.32 和-4.33,Solvation均为1.47,未发生变化。
2.5 mtDNA-ND2基因单倍型分析
共获得13个单倍型,各单倍型在各品种中分布见表1。
由表1可知,H2和H1为优势单倍型,分别占58.3%和13.2%。绵羊群体单倍型多样度(Hd)为0.641,核苷酸多样度(Pi)为:0.357%,平均核苷酸差异数(k)为3.4。塔什库尔干羊、和田羊和多浪羊核苷酸多样度分别为0.330%、0.135%、0.463%。
2.6 绵羊系统进化分析
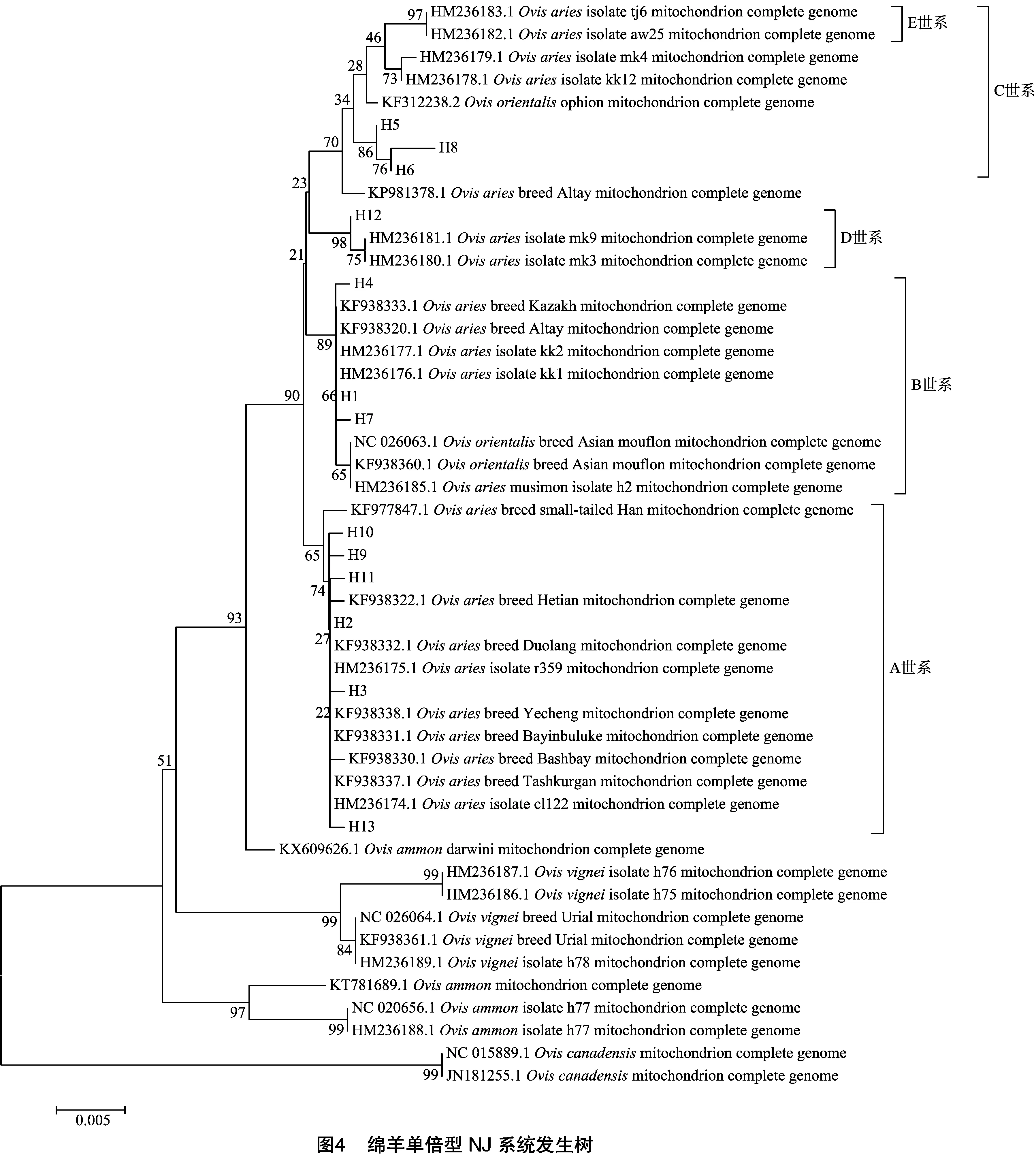
利用MEGA 5.05软件构建绵羊ND2基因不同单倍型NJ系统发育树(图4)。其中包括GenBank下载的36个序列分别是A世系(HM236174-5)、B世系(AM236176-7)、C世系(AM236178-9)、D世系(HM236180-1)、E世系(HM236182-3)序列、盘羊线粒体序列(藏盘羊JX101654.1、帕米尔盘羊KT781689.1、Darwini盘羊KX609626.1、东方盘羊NC020656.1、HM236188.1、加拿大盘羊NC015889.1、JN181255.1)、Vignei(HM236186.1、HM236187.1、HM236189.1、NC026064.1、KF938361)、摩弗伦羊(HM236185、KF312238.1、KF938360.1、NC026063.1)、新疆地方绵羊阿勒泰羊(KF938320.1)、和田羊(KF938322.1)、吐鲁番黑羊(KF938324.1)、巴什拜羊(KF938330.1)巴音布鲁克羊(KF938331.1)、多浪羊(KF938332.1)、哈萨克羊(KF938333.1)、塔什库尔干羊(KF938337.1)、叶城羊(KF938338.1)。

表1 各单倍型在各品种中的分布
由图4可知,单倍型H2、H3、H9、H10、H11和H13属于A世系;单倍型H1、H4和H7与欧洲摩弗仑羊和亚洲摩弗伦羊属于B世系;H5、H6和H8与塞浦路斯摩弗伦羊属于C世系;H12属D世系。
2.7 3个品种间的遗传距离及系统发生树
利用MEGA 5.0计算不同品种间遗传距离(表2),根据遗传距离构建不同品种间NJ系统发生树(图5)。由图5可知,塔什库尔干羊与和田羊关系较近,其次是多浪羊。

表2 3个品种间的遗传距离

3 讨论
绵羊的A+T含量为64.2%,C+G含量为35.8%,A+T含量高于C+G含量;表明绵羊mtDNA-ND2基因含有大量的碱基A和T,存在一定的碱基偏倚性,与笔者前期研究结果[9-10]一致。
基因变异发生在密码子第1、2、3位置分别占9.09%、4.55%、86.36%,与前期研究结果[9]一致。
转换颠换比78.38远大于36.81[9]和转换颠换比的临界值2,说明新疆南疆地区地方绵羊ND2基因序列突变可能未达到饱和状态,转换随着遗传变异增加趋于饱和。
马丽娜等在蒙古羊和小尾寒羊上发现770位点上发生了T到C的转换[11],本研究未发现此变异。
438、504、837、873位点核酸变异只出现在塔什库尔干羊个体中,单倍型H3、H4、H10和H13也只分布于塔什库尔干羊中,是否与其NADH脱氢酶功能差异和低氧适应有联系,仍需进一步研究。另外单倍型H8序列在299位点和883位点均发生突变。突变后ND2亚基空间结构差异不大,但是多肽部分性质发生改变,是否影响线粒体呼吸功能有待进一步探讨。
单倍型H12只分布于多浪羊中,与D世系聚在一起,表明多浪羊存在世系D,与基于细胞色素b的分析结果[9]一致。和田羊未发现世系C,可能与研究样品数量偏少有关。
3个品种平均核苷酸多样度(Pi)为0.357%,塔什库尔干羊、和田羊和多浪羊核苷酸多样度分别为0.330%、0.135%、0.463%,低于赵倩君等[12]、王昕等[13]、张传生等[14]基于细胞色素b研究的0.805%、0.602%、0.850%。但与笔者基于细胞色素C氧化酶亚基Ⅰ(COⅠ)基因分析塔什库尔干羊的0.421%接近。
基于ND2基因的系统发生树表明摩弗伦羊可能是家绵羊世系B的野生祖先,与前人研究观点[5]一致。但是塞浦路斯摩弗伦羊与是世系C聚在一起,可能对世系C有遗传贡献,此观点有待进一步验证。另外塔什库尔干羊与和田羊关系较近,其次是多浪羊,此观点与汤存伟等基于微卫星的分析结果[15]一致。
