陕西榆林东方蜜蜂微卫星DNA种群遗传分析
2018-08-01陈道印刘新宇高宝国周姝婧朱翔杰徐新建郝璐楠周冰峰
陈道印 刘新宇 高宝国 周姝婧 朱翔杰 徐新建 郝璐楠 周冰峰
(1 福建农林大学蜂学学院,福州 350002;2 陕西榆林市草原工作站,榆林 719000;3 陕西榆林市种蜂场,榆林 719000)
蜜蜂是自然界以及农业生产中非常重要的授粉昆虫,具有重大的生态价值和经济价值[1-3]。榆林市位于我国陕西省北部,属于典型的黄土高原地貌,是我国重要的刺槐、枣花以及荞麦蜜源区[4]。上世纪80年代,全国东方蜜蜂资源调查显示我国东方蜜蜂在陕西省北部主要分布于长城以南,榆林市是东方蜜蜂分布的边缘区之一[5]。我们曾利用10个微卫星位点对榆林市的靖边、子洲东方蜜蜂进行遗传分析,研究分布边缘区域东方蜜蜂的遗传多样性和遗传分化,发现靖边的东方蜜蜂与黄土高原其他地区存在种群遗传分化,靖边、子洲的东方蜜蜂微卫星遗传多样性水平低于黄土高原其他地区[6]。靖边东方蜜蜂曾于2009年8月初爆发蜜蜂大量死亡事件[7,8],靖边蜂农所饲养的东方蜜蜂中有近半蜂群死亡[8],种群数量的下降往往会带来遗传多样性的丧失,威胁种群的生存。
微卫星标记(microsatellite)又被称为简单重复序列(SSR),因其具有数量多、变异丰富、容易检测等特点[9],已被广泛应用于种群遗传研究。前人应用微卫星遗传标记对秦巴山区[10]、长白山[11]、黄土高原[6]、海南[12]、浙江[13]等地区的东方蜜蜂进行了种群遗传分化和遗传多样性研究。以前由于受分子生物学实验技术的限制,东方蜜蜂微卫星种群遗传分析所使用的微卫星位点较少,只有5~10个。随着分子生物学实验技术的发展,增加微卫星位点数量对研究东方蜜蜂种群遗传既可行也必要。
本文利用38个微卫星遗传标记对陕西省榆林市东方蜜蜂进行种群遗传分析,评估遗传多样性水平。探索分布在边缘区域的东方蜜蜂在黄土高原生境条件下的遗传分化规律和遗传多样性科学规律,为榆林东方蜜蜂遗传资源的保护和利用奠定理论基础。
1 材料与方法
1.1 样本采集
2017年采集榆林市共7个样点,390群东方蜜蜂(图1)。神木是我们采集的最北部样点,只采集到5群蜜蜂的样本,均为野生蜂群。其余6个样点采自不受人为选育影响的原始饲养蜂群,样本量分别为榆阳43群,佳县66群,吴堡31群,绥德83群,清涧79群,靖边83群。

图1 榆林市各样点分布示意图
1.2 DNA提取与目标片段扩增
每个蜂群提取1只工蜂的基因组,使用Ezup柱式基因组提取试剂盒[生工生物工程(上海)股份有限公司],每个样本均使用38对SSR引物(Ac-2、Ac-5、Ac-26、Ap085、AT101、Ac045、K1458、AT165、UN117、AP208、SV039、BI314、K0715、SV220、AP243、BI278、AP066、SV066、AP148、AP042、AT109、BI216、SV261、AT185、AP249、AC139、UN373、AC011、AT103、AP189、UNEV2、BI225、UN270、244T、Ap313、Ac-1、Ac-35、AT004)进行PCR扩增[14,15]。PCR产物经ABI 3730xl自动测序仪进行基因分型与测序,获得SSR基因型数据。
1.3 数据分析
评估2017年所采集样本的遗传分化状况:Genepop on web分析2017年新采集的各样点之间的遗传分化系数(Fst)[16],并基于Fst计算基因流参数Nm,Nm=1/4(1/Fst-1)[17];Structure 2.3.4进行遗传结构分析[18],生成的结果借助Structure hawester计算最佳分化K值;使用R 3.3.2软件包使用样点作为先验分组进行主成分判别分析(DAPC)验证分化结果[19]。
评估所采集的所有样本的种群遗传多样性水平:Excel Micro-Satellite Toolkit 3.1进行杂合度(He,Ho)、等位基因数(Na)以及多态信息含量(PIC)的计算[20];PopGene 1.31软件进行有效等位基因数(Ne)、香农指数(I)的计算[21];Fstat 3.2软件计算等位基因丰富度(Rs)与近交固定系数(Fis)[22]。
统计所有样本的遗传结构:GenAlex 6.503进行等位基因型以及等位基因频率的统计[23],神木样点由于样本数量过少并未纳入统计。
2 结果
2.1 榆林东方蜜蜂遗传分化
根据样点间的遗传分化系数、Structure分析、DAPC分析等研究,发现榆林东方蜜蜂分化为3个种群,分别为靖边种群、绥德-清涧种群以及佳县-吴堡-榆阳-神木种群。
在Structure分析中,最佳K值为3(图2),靖边样点为独立的一个分组,绥德、清涧被分为一组,神木、榆阳、佳县、吴堡4个样点被分为一组(图3)。
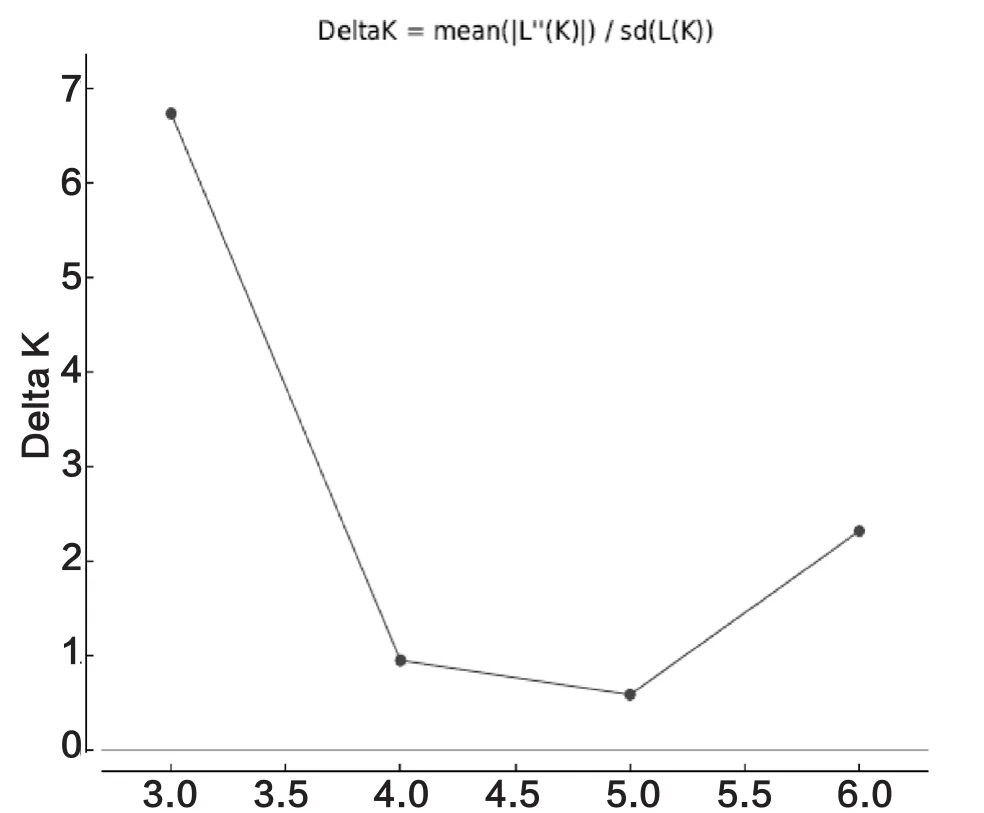
图2 榆林东方蜜蜂Structure最佳分组ΔK曲线图

图3 榆林东方蜜蜂微卫星遗传结构

图4 基于38个微卫星位点的榆林东方蜜蜂DAPC结果图
使用DAPC分析按样点进行分组检验,结果显示靖边为独立的一组,佳县-吴堡-榆阳-神木为一组,绥德-清涧完全重叠为一组(图4)。
遗传分化系数(Fst)以及基因流(Nm)的分析结果(表1)表明,靖边与神木、榆阳、佳县、吴堡之间的Fst在0.0539~0.1083之间。根据Wright所提出的当Fst>0.05时表明两个种群存在遗传分化的标准[24],靖边与神木、榆阳、佳县、吴堡之间存在着明显的分化。
2.2 榆林东方蜜蜂的遗传多样性
遗传多样性参数分析显示榆林东方蜜蜂南部样点的遗传多样性普遍高于北部样点,分布于榆林东部的6个样点在期望杂合度(He)上由南到北逐渐降低,而位于南部种群的绥德、清涧样点在有效等位基因数量(Ne)、香农指数(I)、等位基因丰富度(Rs)以及多态信息含量(PIC)上要明显高于北部种群各个样点。
比较该区域各样点的观察杂合度(Ho)与期望杂合度(He),绥德、清涧、靖边、吴堡以及榆阳5个样点的观察杂合度(Ho)低于期望杂合度(He),近交系数(F-is)均大于0。
2.3 榆林东方蜜蜂的微卫星遗传结构
榆林市各个种群的东方蜜蜂在9个微卫星位点上表现出不同的遗传结构。在位点Ap085上,佳县与吴堡样点主要以a187(0.389~0.409)为优势等位基因,而其他样点等位基因分布较为分散。在位点AT101上,佳县、吴堡与榆阳以a246(0.5~0.583)为优势等位基因,靖边、绥德、清涧则以a244(0.259~0.462)、a245(0.231~0.398)为优势等位基因。在位点AC045上,绥德、清涧拥有相对较高的a142比例(0.120~0.175)。在位点AP243中,佳县、吴堡、榆阳以a257为优势等位基因(0.326~0.492),其余样点的等位基因比较分散并无优势等位基因。在SV066中,靖边的a185占比在0.97左右,其余样点则拥有相对较高的a180比例(0.105~0.241)。在位点AP042中,绥德、清涧的a139(0.095~0.211)拥有相对较高的频率,其余样点几乎仅以a143为唯一等位基因。在位点AC011中,榆阳、佳县、吴堡拥有相对较低的a134(0~0.038),其余样点的a134在0.108~0.171之间。在位点AP189中,靖边以a120(0.367)、a122(0.349)为优势等位基因,其余样点则以a118(0.339~0.535)为优势等位基因。在位点244T中,靖边主要以a189(0.228)、a197(0.321)为优势等位基因,其余样点则以a189(0.171~0.417)、a191(0.169~0.317)、a193(0.037~0.274)为优势等位基因(表3)。

表1 榆林东方蜜蜂遗传分化系数与基因流

表2 榆林东方蜜蜂遗传多样性水平参数信息
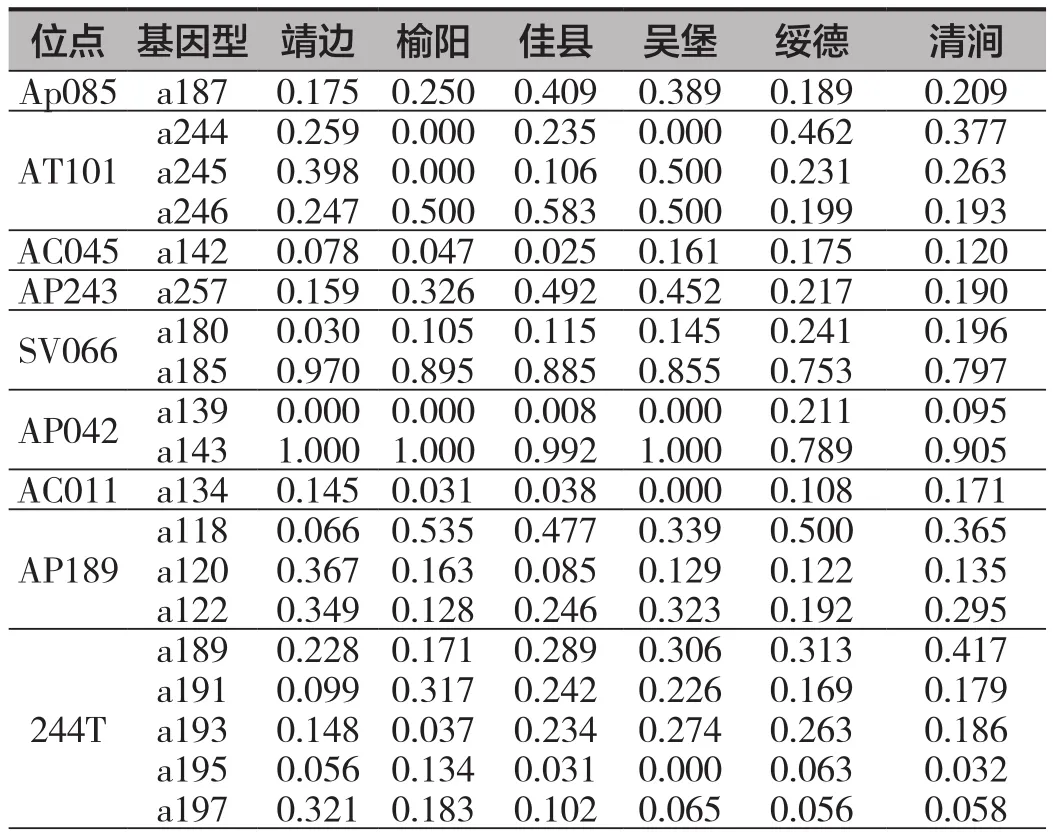
表3 榆林东方蜜蜂样点之间存在差异位点的等位基因频率表
3 讨论
3.1 榆林东方蜜蜂的遗传多样性水平
遗传多样性是物种进化的基础[25],因此遗传多样性评估对于遗传资源的鉴定以及保护有着非常重要的意义。榆林市南部东方蜜蜂种群的遗传多样性普遍高于北部种群的多样性水平。利用本研究与其他研究中相同的8个SSR位点(Ac1、Ac2、Ac5、Ac26、Ac35、Ap085、A313、AT101)进行期望杂合度的计算与比对,海南的平均期望杂合度为0.5970[12],黄土高原其他样点的平均期望杂合度为0.6781[6],秦巴山区的平均期望杂合度为0.6877[10],榆林市的平均期望杂合度为0.6297,表明榆林市东方蜜蜂遗传多样性水平在黄土高原较低,也低于秦巴山区东方蜜蜂的遗传多样性水平。
榆林市植被分布由南到北逐渐稀疏,仅于部分沟壑中可能存在有树洞的乔木,这意味着榆林野生东方蜜蜂的营巢条件受到限制,影响当地东方蜜蜂的自然种群数量,使得榆林东方蜜蜂的遗传多样性呈现较低水平。生境破碎化造成东方蜜蜂分布的破碎化,影响各种群间的基因交流,使种群内的东方蜜蜂存在一定程度的近交现象,丧失部分遗传多样性。有观点认为中蜂囊状幼虫病能够导致东方蜜蜂在某些位点上的等位基因大量丢失,造成遗传多样性的降低[10]。据采样时了解到,榆林市部分县市曾爆发中蜂囊状幼虫病,这可能也是造成榆林市东方蜜蜂种群遗传多样性较低的原因之一。
3.2 榆林东方蜜蜂的遗传分化机制
榆林市属于典型的黄土高原地貌,气候干燥,植被稀疏,生态环境极为脆弱[26],境内生境破碎化严重[27]。生境破碎影响种群分布,阻碍种群间基因流,促使种群发生分化[28]。榆林所发现的3个种群之间直线距离均在30km以上,蜜蜂最远的婚飞距离是15km[29],而各个种群之间缺乏野生东方蜜蜂营巢条件,也没有蜂场,从而限制了各个种群之间东方蜜蜂的迁移,从而阻断了种群间的基因流,造成各个种群间发生种群遗传分化。
