微生物碳酸盐岩储层特征及主控因素
——以塔里木盆地阿克苏地区下寒武统肖尔布拉克组上段为例
2018-07-30翟秀芬周川闽
白 莹 罗 平 刘 伟 翟秀芬 周川闽
( 中国石油勘探开发研究院 )
Microbialite一词首先由Burne和Moore提出[1],后经Riding改为Microbolite[2],特指由微生物形成或与微生物有关的岩石[2-6],在中国这种多具特殊微生物沉积构造的岩石被称为微生物岩[7-13]。目前研究最多、分布最广的微生物岩为微生物碳酸盐岩,与之有关的微生物多为蓝细菌,主要包括葛万菌、肾状菌和表附菌等[2-6,14-17]。微生物碳酸盐岩主要集中分布在中—新元古代、寒武纪和早奥陶世这类埋藏较深的古老地层中,目前国内外微生物碳酸盐岩已经获得了重大油气发现,如美国阿拉巴马州阿普尔顿油田和东西伯利亚里菲系油气田,以及中国渤海湾盆地华北油田和四川盆地威远气田、资阳气藏,表明其可形成良好的储层[7,11-13,18-21]。
近年来,塔里木盆地盐下微生物碳酸盐岩成为了油气勘探的热点,比如中深5井在寒武系台内微生物岩地层中产出了工业气流,中深1井寒武系肖尔布拉克组取得了重大油气突破。这些发现直接使塔里木盆地下寒武统肖上段微生物岩成为了研究重点,然而相关的储层特征研究略显不足[13,22-27]。本文基于野外剖面详测和室内薄片观察,对塔里木盆地阿克苏地区下寒武统肖上段微生物碳酸盐岩的岩石结构进行研究,并对不同类型的微生物碳酸盐岩储层进行分类和描述,探讨储层主控因素,力求更加系统地认识微生物碳酸盐岩储层,为塔里木盆地下寒武统的油气勘探提供理论依据。
1 研究区地质背景
阿克苏露头区位于新疆维吾尔自治区阿克苏市境内,隶属塔北柯坪断隆东段,在早寒武世其大致位于塔西台地台缘带附近(图1)[23-25,28]。本文自西向东选取了6个剖面进行研究,依次为于提希、苏盖特布拉克、磷矿沟、肖尔布拉克、什艾日克、库勒剖面。其中于提希和苏盖特布拉克剖面位于台缘带,其他剖面位于台内带(图2)。
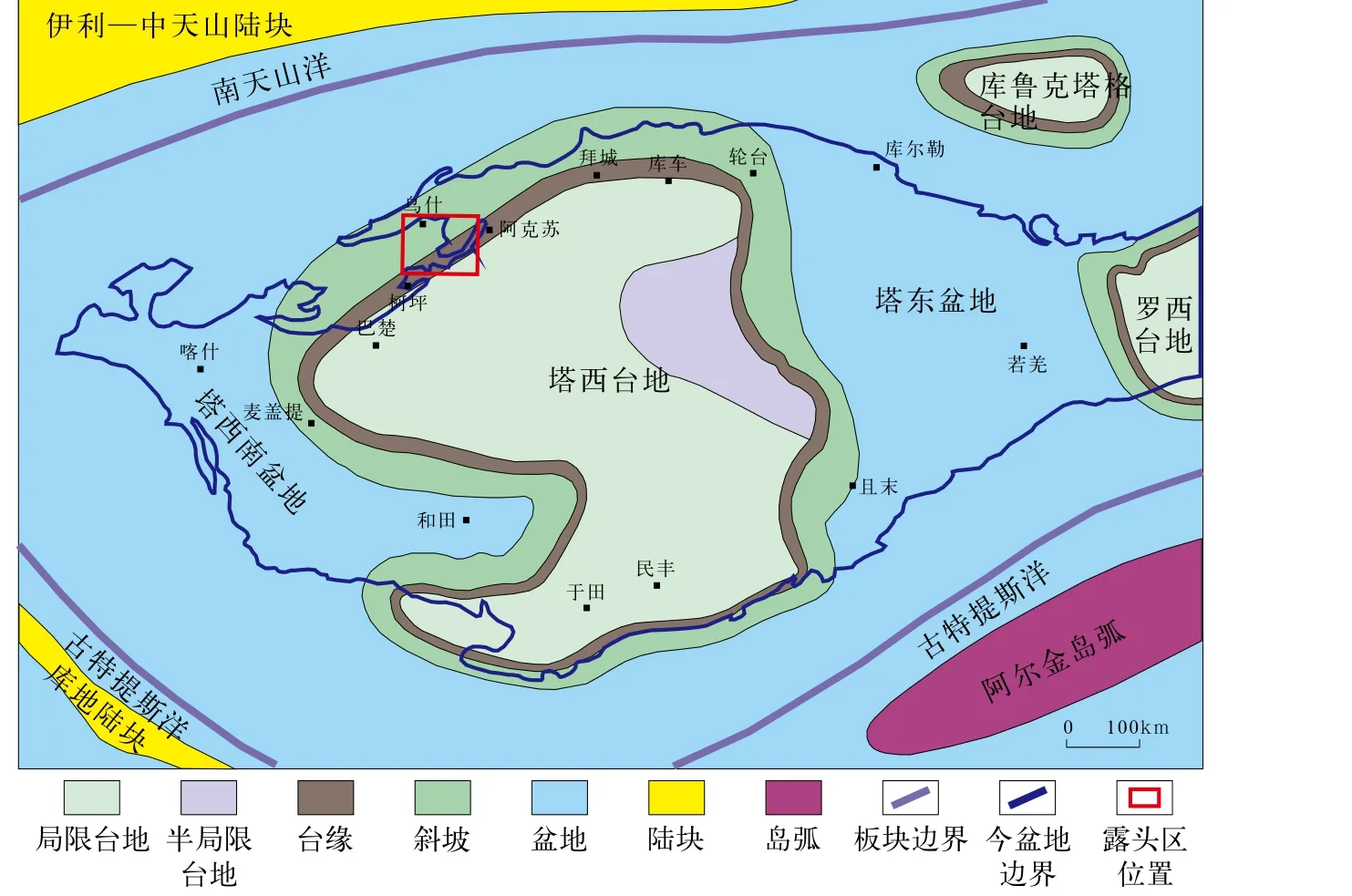
图1 塔里木盆地阿克苏地区早寒武世岩相古地理图[28]

图2 塔里木盆地阿克苏地区剖面位置图
新元古代末期塔里木板块的边缘开始裂解,从南华纪至寒武纪依次经历了断陷阶段、坳陷阶段和广海碳酸盐台地阶段,早寒武世塔里木板块处于赤道附近的低纬度地区,大概位于冈瓦纳古陆西部[29-32]。以塔北地区为例,早寒武世早期发生的大规模海侵事件使得不整合面上沉积了由黑色页岩、硅质岩和磷质岩构成的下寒武统玉尔吐斯组[23-25,30-32]。上覆的下寒武统肖尔布拉克组可分为两段,其中下段厚约30~40m,代表岩性为深灰色层状孔洞球粒白云岩,指示水体较深的碳酸盐缓坡;上段厚约80~100m,主要发育微生物建造,缓坡上的进积型礁滩体组成了新台缘,标志着碳酸盐缓坡正式演变为镶边碳酸盐台地(图3)。
2 微生物碳酸盐岩岩石结构特征
阿克苏露头区肖上段主要发育了两种尺度范围为米级至千米级的微生物巨观建造,其中微生物礁建造主要位于台缘带,而微生物层建造可位于台缘带和台内带。宏观上,微生物礁建造可进一步分为凝块石礁和表附菌礁;微生物层建造可分为凝块石层、表附菌层、叠层石层、泡沫绵层和层纹石层(图4)。
凝块石礁(图4a-e)主要发育在苏盖特布拉克剖面和于提希剖面,是小型台缘微生物礁的主体部分。凝块主要由泥晶组成,大小约(0.2~3)cm×(0.3~3)cm,之间以亮晶胶结。凝块分布不均,大小不一,主要包括树枝状(图5a、b)和团块状(图5c、d)这两种中观形态,其中前者主要由微生物碎屑及砂屑组成,包壳一般由暗色泥晶组成,是由微生物参与形成的[8,33-34],可见由泥晶胶结而成的馕状—椭圆状肾形菌,直径约0.1~1cm(图5e),这种肾形菌的形态和组合方式同样可见于国内的四川盆地震旦系灯影组,以及西澳大利亚和比利时地区上泥盆统和加拿大纽芬兰地区下奥陶统[35-36];后者主要由聚集成团的包壳颗粒和砂屑组成,单个包壳颗粒及砂屑直径约0.1~1mm,集合体直径约1~5mm(图5f)。
表附菌礁主要发育在苏盖特布拉克剖面,是大型台缘微生物礁的主体部分(图4a、5g)。表附菌礁中观上可由斑状凝块组成(图5h)。微观上树枝状表附菌集合体呈灌木状,长约1~7mm,宽约1~5mm,与鲁西张夏组微生物岩中发现的灌木状表附菌类似[11-12,37],主要由相对较长的具分叉的叶状体组成。这些向上生长的叶状体主要呈丛状,向上、向外不断生长壮大,单个叶状体长0.1~1mm,宽0.1~0.5mm。表附菌集合体之间发育孔隙,孔隙可被粉晶白云石充填(图5i)。

图3 塔里木盆地阿克苏地区于提希剖面地层柱状图

图4 塔里木盆地阿克苏地区微生物巨观构造分类

图5 塔里木盆地阿克苏地区微生物礁特征
凝块石层分布较广,在典型剖面均有发育。中观上凝块石层厚约3~10cm(图6a、b),层面弯曲,具有亮暗相间的不规则凝块结构,大小约(0.5~1.5)cm×(0.5~3)cm,溶蚀孔洞多顺层分布,大小可从毫米级至厘米级,多呈圆形—椭圆形,溶孔密度约为10~20个/dm2。微观上可见包壳结构,直径约0.2~1cm,其中暗色包壳主要由泥晶白云石构成,形成边缘呈发散状的暗色包边,可能为微生物稳定作用的产物[8,33-34],一般厚约0.1~2mm;浅色的核心由粉晶白云石组成,核心内部发育粒间溶孔和晶间溶孔,多被亮晶自形白云石充填—半充填(图6c)[26]。

图6 塔里木盆地阿克苏地区微生物层特征
表附菌层主要分布在于提希剖面(图6d)和苏盖特布拉克剖面(图4b)。其中于提希剖面的风暴沉积物不甚发育,而在苏盖特布拉克剖面,表附菌层往往和风暴岩伴生。于提希剖面成岩作用较为强烈,宏观上的表附菌层厚5~15cm,层面平直,在中观上主要呈不规则斑状,斑块大小约(0.2~1)cm×(0.4~1)cm,分布无规律(图6e)。微观特征类似于表附菌礁,主要由灌木状的树枝状表附菌集合体组成(图6f)。在微生物结构保存较好的苏盖特布拉克剖面,层面平直的表附菌层厚10~15cm(图6g),中观上呈团块状—斑状,团块/斑块多由房室状凝块组成,凝块直径约0.2~1cm,多集中在0.8~1cm(图6h),房室壁厚0.1~0.2cm,具有中空结构。微观上半闭合—闭合房室状表附菌集合体主要由相对较短的椭圆形叶状体放射排列组成,大小约为(50~100)μm×(100~1000)μm,叶状体之间和房室状集合体中心的孔隙多被胶结物充填(图6i)。这可能说明房室状表附菌和灌木状表附菌相比,前者的水动力生长环境更强[11-12,37]。
叠层石层主要分布在肖尔布拉克剖面和什艾日克剖面。宏观上肖上段主要为层状叠层石(图6j),层厚约20cm;中观上表现为多条相互平行、且横向连续性较好的叠层结构,中观层间距约1cm左右(图6k);微观上叠层结构明暗相间,微观层间距约2mm,其中暗层通常由粉晶—泥晶组成,往往含有较多的泥质及有机质,而亮层多由粉晶—细晶组成(图6l)。
泡沫绵层主要见于什艾日克剖面、磷矿沟剖面和肖尔布拉克剖面。层状的泡沫绵层,中观上为浅灰色厚层状泡沫结构,层厚约5~10cm,借助放大镜可见类似砂屑的颗粒密集排列,可见圆形的针状溶孔,直径约1~3mm(图6m)。微观上可见海绵状骨架,一般由多个圆球状至椭球状的泡沫状蓝细菌以点接触方式的排列组成,在塔西北震旦系也有发育[38-39]。单个泡沫状蓝细菌包括泥晶边缘和亮晶空腔,大小一般为0.2~1mm(图6n)。在什艾日克剖面和磷矿沟剖面,可见圆球状的泡沫状蓝细菌构成平行排列的蜂窝状泡沫绵层(图6o),而在肖尔布拉克剖面,椭球状的泡沫状蓝细菌组成了杂乱排列的不规则状泡沫绵层。
层纹石层主要分布在于提希剖面和磷矿沟剖面。层状层纹石层可见平行纹层结构,层间距为1~2mm,有时横向连续性不好,呈断续分布(图6p、q)。微观上层纹石层可由菌纹层、颗粒纹层和亮层组成(图6r)。菌纹层由多条细密的有机质纹层组成,单条有机质纹层厚10~50μm,集合体宽约1~2mm。颗粒纹层多由包壳颗粒和蓝细菌碎屑组成,粒级多集中在100~500μm。亮层则为暗层之间的胶结物。其中菌纹层和颗粒纹层可看做是暗层,但与叠层石层相比,层纹石层亮暗层的宽度和间隔较大,且含有较多的砂屑、包壳颗粒以及微生物成分[40]。
上述7种微生物岩结构中,凝块石礁、凝块石层、叠层石层、层纹石层与微生物以及微生物席的黏结和捕集作用有关,而表附菌礁、表附菌层和泡沫绵层与钙质微生物群落本身生长和自身格架有关。
3 微生物碳酸盐岩储层特征
3.1 储层类型
对阿克苏露头区下寒武统肖上段微生物碳酸盐岩的岩石结构数量进行统计,结果显示肖上段微生物储层以凝块石礁储层和泡沫绵层储层为主。
3.2 孔隙类型
扫描电镜、显微镜以及岩石抛光面的观察结果显示,肖尔布拉克组微生物礁具有类型丰富的微生物相关孔隙,包括窗格孔、微生物组构架间溶孔、微生物组构腔内溶孔、微生物组构铸模孔、微生物组构超大溶孔以及与泡沫绵层相关的孔隙等,同时也具有普通碳酸盐岩储层所具有的非微生物结构储集空间。
对于与微生物相关的孔隙来说,窗格孔的形成可能和微生物的活动、生物腐烂以及成岩时的干涸收缩有关[41],直径多为0.3~1cm,可被亮晶白云石全充填或半充填,局部可能残留晶间孔,一般认为窗格孔多发育在具有纹层状结构的微生物岩中(图7a)[23,38-39]。在阿克苏露头区,直径约2mm的孤立、不规则窗格孔可见于于提希剖面的层纹石或者与之相关的颗粒岩中。微观下可见呈菱形的窗格孔紧邻层纹石层的菌纹层发育,说明其可能为微生物成因(图7b)。
微生物组构架间溶孔位于微骨架之间,包括亮晶胶结物中的溶孔,以及少量未被亮晶胶结物充填的原生孔隙。比如在于提希剖面的凝块石礁中,孤立的架间溶孔主要存在于凝块骨架之间的胶结物中,可为大小约(1~5)mm×(1~2)mm的不规则溶孔至直径小于1mm的针孔状溶孔(图7c、d)。在苏盖特布拉克剖面的表附菌层中,由房室状表附菌集合体组成的骨架、架间以及集合体内部多被亮晶—粉晶白云石充填,溶蚀孔洞多孤立分布于架间(图5e)。在于提希剖面的表附菌层中,树枝状表附菌骨架之间的胶结物内部可见圆形溶孔成群分布,连通性不好,直径多为200μm(图7f)。
微生物组构腔内溶孔是指微生物组构腔内发育的溶蚀孔隙以及少量的原生孔隙,前提是微生物的残余腔壁仍有保留。比如在阿克苏露头区广泛发育的包壳凝块石层建造,暗色泥晶包壳内部的粉晶内核多被溶蚀,形成以腔内晶间溶孔为主的孤立孔隙,孔隙形状多和包壳凝块的整体形状有关,直径一般为0.5~1mm(图7g)。

图7 塔里木盆地阿克苏地区微生物储层孔隙特征
微生物组构铸模孔的成因类似于腔内溶孔,只是溶蚀程度更高一些,是微生物组构被完全溶蚀后的产物,仅仅保留了组构的形状。微生物组构铸模孔多存在于层纹石层的颗粒层中,或与之伴生的颗粒岩中,部分可见微生物碎屑残留的铸模孔,多为圆形,直径多集中在500μm左右(图7h)。
微生物组构超大溶孔是指溶蚀孔隙大于微生物组构本身的一类储集空间,不具有固定的形状,但一般为毫米级,部分可达厘米级,大多数为微生物组构架间溶孔、腔内溶孔、铸模孔或普通储集空间后期又经历了表生岩溶作用而形成的(图7i)。
缝合线(图7j、k)和微裂缝(图7l)是比较常见的非微生物结构储集空间。缝合线一般和微生物岩本身的沉积环境、埋藏深度以及矿物学应力方向等因素有关。微裂缝则有利于增加孔隙连通性,并为孔隙的进一步溶蚀和发育创造条件[23-26,41]。
在阿克苏露头区,泡沫绵层中的泡沫可呈平行排列状,被称为泡沫绵层叠层石(图8a),亦可呈不规则状排列,被称为非叠层状泡沫绵层(图8b)[23-26,38-39]。与泡沫绵层相关的孔隙包括泡沫绵层腔内溶孔、泡沫绵层腔铸模孔和泡沫绵层超大溶孔。泡沫绵层腔内溶孔普遍发育,位于单个泡沫内,绝大部分亮晶层腔和泥晶边缘基本未被溶蚀,因此孔隙多为孤立发育,连通性不好(图8c、d)。泡沫绵层腔铸模孔发育在保留残余泥晶边缘的泡沫内,有时可见少量的残余亮晶层腔(图8e)。铸模孔主要发育在单个泡沫内,具有一定的连通性,发育较为广泛,是泡沫绵层白云岩中较为重要的储集空间。泡沫绵层超大溶孔为单个泡沫被彻底溶蚀后,所产生的大于泡沫本身的溶蚀孔隙,主要为顺层分布,连通性较好,这类孔隙发育广泛,是泡沫绵层白云岩中最重要的储集空间(图8f)。

图8 塔里木盆地阿克苏地区与泡沫绵层有关的孔隙特征
3.3 储层物性特征
总体来说微生物白云岩储层的孔隙度一般分布在1%~10%之间,最低只有0.4%,最高可达15.38%。但是渗透率普遍较低,多为0.1mD左右,可低至0.07mD,最高可达37.351mD(图9a);过高的渗透率样品有3个,可能是受到了地表风化作用的影响。铸体薄片观察结果也显示该地区储层的孔隙连通性普遍较差,基本属于中孔、低—特低渗透储层。

图9 塔里木盆地阿克苏地区储层物性特征
通过镜下观察以及物性统计,综合考虑微生物岩储层的孔隙特征及连通情况,认为凝块石礁储层和泡沫绵层储层为较有利的储集岩石类型,而其他类型的微生物岩则物性较差。其中凝块石礁储层的实测孔隙度大多介于2%~10%之间,最低至0.52%,渗透率跨度较大,具有两个峰值,比如在于提希剖面一般为1mD左右,属于低孔低渗透储层,而在苏盖特布拉克剖面,大部分样品的渗透率可达15~40mD,可归为低—中孔、低—中渗透储层(图9b、c)。而泡沫绵层主要发育在什艾日克剖面和肖尔布拉克剖面,两个剖面物性特征类似,孔隙度均介于3.2%~8.38%,渗透率均介于0.010~1.537mD,绝大多数在1mD以上,属于低孔低渗透储层。这说明不同类型的微生物岩储层具有较大的物性差别,相同类型的微生物岩储层在不同的剖面也具有较大的区别。
4 微生物碳酸盐岩储层主控因素
和碎屑岩储层相比,碳酸盐岩储层的形成除了严格受控于沉积作用和构造作用之外,与成岩作用更加密不可分。然而在微生物碳酸盐岩储层中,微生物结构对储集空间结构的控制作用也不可忽视。
4.1 沉积作用
前人的研究成果指出,在塔里木盆地下寒武统中,沉积相决定了微生物碳酸盐岩储层的基本类型和展布特征[23-25]。肖上段沉积时期处于碳酸盐缓坡至镶边碳酸盐台地的转变期,阿克苏露头区可分为台缘带和台内带,这两个不同的相带发育不同的微生物结构。例如,纹层、叠层和泡沫绵层等层状结构主要发育在台内带微生物碳酸盐岩中,而具有凝块结构的凝块石礁主要发育在台缘带,具有凝块结构的凝块石层可发育在台缘带或者台内带。
4.2 古地貌特征
震旦纪末期柯坪运动导致了高低不平的古地貌的形成,古高地、古斜坡与古洼地的存在控制着微生物碳酸盐岩储层的展布以及成岩方式[23-25]。比如台内带的泡沫绵层和凝块石层倾向于发育在古高地,较高的地势更容易受到淡水淋滤等溶蚀作用,溶孔也更为发育;台缘带的凝块石礁倾向于发育在古斜坡处,溶蚀作用相对较弱,溶孔发育程度较弱。
4.3 成岩作用
4.3.1 埋藏压实与胶结作用
露头(图10a)、镜下(图7j、k)可见微生物碳酸盐岩中多发育压溶缝合线,同时层纹石中的砂屑、包壳颗粒以及微生物成分有破碎的现象,部分具有定向分布和压扁的特征(图7h),这说明肖上段曾经经历过强烈的压实作用。准同生期或沉积期形成的原生孔隙,比如窗格孔、微生物组构架间溶孔(原生)、微生物组构腔内溶孔(原生)、泡沫绵层腔内溶孔等已经被亮晶胶结物充填(图7、图8)。因此在埋藏压实与胶结的综合作用下,早期形成的原生孔隙基本消失殆尽。然而从储层改造方面来看,经过埋藏压实作用产生的缝合线和微裂缝很容易遭受溶蚀并形成新的储集空间,也可以将孤立的孔隙连通,改善储层质量(图 7i—l)[23-26,38-39,42]。

图10 塔里木盆地阿克苏地区微生物储层特征
4.3.2 溶蚀作用
溶蚀作用按流体性质可分为大气水溶蚀作用、热液溶蚀作用和有机酸溶蚀作用,它们可以改善微生物碳酸盐岩储层的储集性能,因此对微生物碳酸盐岩储层的形成至关重要。
对阿克苏露头区来说,大气水溶蚀作用对储层的影响最大。其中腔内溶孔和铸模孔受大气水的选择性溶蚀作用影响,主要产生于准同生期或同生期未固结成岩的颗粒内部(图7g、h)。至表生期,大气水的溶蚀作用则不具选择性,微生物岩中可见由微裂缝和缝合线经淋滤、溶解作用形成的褐铁矿化的压溶缝合线(图10b、c),它们可作为大气淡水的溶蚀通道,将先前形成的孔隙扩大、连通,并形成超大溶孔[38-39,41](图7i)。
热液溶蚀作用和有机酸溶蚀作用对储层的影响,前人已经有较为详细的研究。比如宋金民等认为阿克苏地区的斑马纹白云岩为热液成因的典型代表,并指出沿裂缝和断裂上涌的深部热液可将围岩溶蚀并产生孔隙[25];张静等指出柯坪地区寒武系—奥陶系剖面可见大量热液成因的粗晶孔洞白云岩,说明塔里木盆地具有发育热液白云岩储层的条件[43];随着埋藏深度和温度的增加,具有较高有机质含量的微生物岩可产生有机酸、CO2等酸性物质,贾承造等和胡明毅亦指出奥陶纪为玉尔吐斯组黑色页岩的生油高峰,油气倾向于运移到上覆寒武系微生物白云岩中,可产生有机酸溶蚀作用,会对岩石本身进行溶蚀并形成孔隙[29,44]。
在于提希剖面的凝块石礁中发现了与凝块石伴生的斑马纹白云石(图10d),镜下可见鞍状白云石多发育溶蚀孔洞,这可能是由于相关热液中的Mg2+较少,而Ca2+较多,因此鞍状白云石不甚稳定,易被溶蚀改造(图10e)。在苏盖特布拉克剖面,可见礁岩中有沥青充填,说明其曾经经历过油气的运移及充注(图10f)[23]。然而这些证据也只能证明热液溶蚀作用和有机酸溶蚀作用可能有利于溶孔的形成,进一步的证据仍需要补充。不过可以明确的是这两种溶蚀作用的规模和影响远不及大气水溶蚀作用。
4.4 微生物结构
具有微生物结构的微生物碳酸盐岩具有不同的原生孔隙类型与结构特征,这是由于不同类型的微生物之间结构相差很大的缘故。对于次生孔隙来说,微生物结构对其形成具有一定的保护作用,这是因为微生物结构的钙化成岩时间较早,一般不会晚于成岩早期,所形成的坚硬格架有利于后期溶孔的形成与保存,而且不同类型的格架对溶孔的保存影响不一。孔隙度较差的宏观微生物岩建造包括表附菌礁、表附菌层、叠层石层和层纹石层。其中表附菌礁多为由毫米—厘米级表附菌树枝状集合体组成的微型骨架,孤立溶孔多位于二分叉叶状体之间(图5i);表附菌层多为由毫米—厘米级表附菌房室状/灌木丛状集合体组成的微型骨架,孤立溶孔多位于房室内部及边缘(图6f、 i);叠层石层和层纹石层可发育窗格孔及顺层溶孔,然而成岩早期发育的窗格孔多被胶结,而顺层孔洞穿层沟通性不好,因此整体物性较差(图7a)。孔隙度较好的宏观微生物岩建造包括凝块石礁、凝块石层和泡沫绵层。其中凝块石礁可具厘米—分米级树枝状—团块状骨架,以发育凝块间溶孔为特征(图7d);凝块石层多具有包壳结构,溶孔多位于包壳颗粒内部以及包壳颗粒的边缘,强烈的溶蚀作用以及破裂作用使得孔隙沟通性较好(图7g);泡沫绵层的原生孔隙一般位于单个泡沫腔内,并以此为基础形成了与泡沫绵层相关的孔隙,包括泡沫绵层腔内溶孔、泡沫绵层腔铸模孔和泡沫绵层超大溶孔,溶孔多趋于顺层发育,上下连通性较好(图8)。
5 结论
(1)在塔里木盆地阿克苏露头区,下寒武统肖尔布拉克组上段主要发育微生物礁和微生物层这两种微生物岩巨观构造,其中前者可分为凝块石礁和表附菌礁,后者可分为凝块石层、表附菌层、叠层石层、层纹石层和泡沫绵层。
(2)微生物碳酸盐岩储层具有与微生物相关孔隙和非微生物结构储集空间,其中前者包括窗格孔、微生物组构架间溶孔、微生物组构腔内溶孔、微生物组构铸模孔、微生物组构超大溶孔以及与泡沫绵层相关的孔隙,后者包括缝合线和微裂缝。
(3)凝块石礁储层在于提希剖面属于低孔低渗透储层;而在苏盖特布拉克剖面属于低—中孔、低—中渗透储层;泡沫绵层储层在什艾日克剖面和肖尔布拉克剖面均属于低孔低渗透储层,这说明不同类型的微生物岩储层具有较大的物性差别,相同类型的微生物岩储层也具有地区差异性。
(4)沉积相决定了微生物碳酸盐岩储层的基本类型和展布特征,古地貌控制着微生物碳酸盐岩储层的展布范围以及成岩方式。成岩作用是微生物碳酸盐岩储集空间形成的关键因素,而微生物结构影响了潜在储集空间的形成。
