杜拉鲁肽对阿尔茨海默病样神经退行性变的保护作用
2018-07-27陈沿霖郭爱彭鹏周梅谷中娅刘筱晗张梦喆邓艳秋
陈沿霖,郭爱,彭鹏,周梅,谷中娅,刘筱晗,张梦喆,邓艳秋
阿尔茨海默病(AD)又称老年性痴呆,是一种常见的以进行性学习记忆减退为主要临床症状的神经退行性疾病,其两大特征性病理改变是细胞间淀粉样蛋白(Aβ)沉积形成的老年斑(SP)和神经细胞内神经原纤维缠结(NFTs),且NFTs与老年痴呆程度呈正相关[1-2]。过度磷酸化的细胞骨架蛋白Tau和神经丝(NFs)与NFTs形成密切相关,是AD的早期病变,其在AD神经变性发生发展中起着非常重要的作用[3]。Tau蛋白和NFs的磷酸化和脑内糖代谢产生的O-糖基化存在竞争关系,AD脑内糖代谢和胰岛素信号紊乱与Tau蛋白和NFs的磷酸化密切相关[4-5]。而2型糖尿病(T2DM)也存在胰岛素信号异常和糖代谢紊乱,研究表明T2DM患者AD发病率增高且发病时间提前,因此AD也被称为第三类型糖尿病[6-7]。胰高血糖素样肽(GLP)-1是一种肠促胰岛素,前期研究发现GLP-1类似物如利拉鲁肽和受体激动剂艾塞那肽能改善AD鼠模型的记忆和认知功能[8-10]。本研究通过应用GLP-1受体(GLP-1R)拮抗剂Ex9-39和PI3K抑制剂渥曼青霉素(wortmannin)观察杜拉鲁肽对人神经母细胞瘤SH-SY5Y细胞AD样变的Tau和NFs异常磷酸化的保护作用并探讨其机制,为改善AD药物的研发提供依据[11]。
1 材料与方法
1.1 材料 SH-SY5Y细胞购自中科院上海细胞库,杜拉鲁肽、PI3K抑制剂wortmannin和GLP-1R拮抗剂Ex9-39分别购自礼来公司、Sigma公司和杭州中肽公司;Tau和神经丝蛋白的抗体购自Thermo Fisher和Biolegend公司;R61d多克隆抗体由美国IBR研究所馈赠;兔源Total/Phospho-PI3K(p85),Total/Phospho-GSK3β(S9)多克隆抗体购自 Cell Signaling Technology;辣根过氧化物酶标记的羊抗小鼠抗体和羊抗兔抗体购自Abcam公司;β-actin抗体、BCA试剂盒、组织裂解液RIPA和ECL化学发光试剂盒分别购自中国碧云天生物技术研究所、索莱宝公司和Milipore公司。
1.2 方法
1.2.1 细胞培养与分组处理 SH-SY5Y细胞采用DMEM/F12培养基(含10%胎牛血清、0.1%链霉素+青霉素)于37℃、5%CO2孵育中贴壁生长,每2~3 d更换1次培养基。细胞密度达到70%~80%时,传代或用于实验。细胞分为6组:空白对照组(CON组),磷酸盐缓冲液(PBS)处理12 h;wortamannin干预组(W组),1µmol/L wortamannin处理12 h;杜拉鲁肽干预组(D组),10 nmol/L杜拉鲁肽处理12 h;杜拉鲁肽与wortmannin共同干预组(D+W组),10 nmol/L杜拉鲁肽和1µmol/L wortamannin共同处理12 h;杜拉鲁肽+wortmannin+Ex9-39共同干预组(D+W+E组),10 nmol/L杜拉鲁肽、1µmol/L wortamannin和10µmol/L Ex9-39共同处理12 h;Ex9-39干预组(E组),10µmol/L Ex9-39处理12 h。各药物浓度根据前期实验和预实验结果选定[12]。
1.2.2 改良MTT法测定细胞活性 胰酶消化SH-SY5Y后室温1 000×g离心5 min,弃去上清,培养基重悬细胞后1×104个/孔接种于96孔板,每组10个复孔。待生长至80%汇合度时,细胞无血清化后按照上述分组处理,处理结束后每孔加入0.5%MTT溶液10µL,37℃孵育4 h后每孔加入100µL DMSO于摇床上轻摇10 min,酶标仪检测490 nm波长处各孔光密度(OD)值。
1.2.3 Western blot检测蛋白表达水平 细胞以1×106个/孔接种于6孔板,参照1.2.1进行分组处理,处理后的细胞加入RIPA裂解液(含1 mmol/L PMSF)4℃裂解30 min后,16 000×g离心10 min取上清。BCA法测定蛋白浓度,十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE,7.5%或10%分离胶),湿转法(280 V,300 mA转膜1.5 h)将蛋白转至PVDF膜上,5%BSA封闭30 min后,一抗4℃孵育过夜,二抗(1∶10 000)室温2 h,ECL发光,曝光,Image J软件分析各检测结果。用β-actin做内参,Tau5抗体检测Tau蛋白总量,tau[pS396]、tau[pS404]、tau[pS262]抗体检测Tau蛋白在396、404和262位点的磷酸化水平;用SMI31抗体检测细胞内中、高分子质量神经丝蛋白(NF-M和NF-H)的磷酸化水平,以识别总神经丝蛋白的R61d为内参,检测NF-M和NF-H的总体水平;用Total/Phospho-PI3K(p85)抗体和 Total/Phospho-GSK3β(S9)抗体检测PI3K和GSK3β的磷酸化水平。
1.3 统计学方法 采用SPSS 22.0统计学软件进行数据分析。符合正态分布的计量资料以均数±标准差(±s)表示,CON组、W组、D组和D+W组均数比较采用析因设计的两因素方差分析,CON组与E组、D+W组与D+W+E组均数比较均采用两独立样本t检验,用Graphpad Prism 5.0进行统计分析和作图,P<0.05为差异有统计学意义。
2 结果
2.1 MTT法检测不同药物对细胞活力的影响 杜拉鲁肽干预后细胞活力上升(F=293.112,P<0.01);wortamannin干预后SH-SY5Y细胞活力下降(FW=162.630,P<0.01),杜拉鲁肽和wortamannin存在交互效应,D+W组细胞活力上升(F交互=19.840,P<0.05)。Ex9-39干预后SH-SY5Y细胞活力下降(t=4.423,P<0.01),D+W+E组的细胞活力较D+W组降低(t=9.406,P<0.01),见图1。
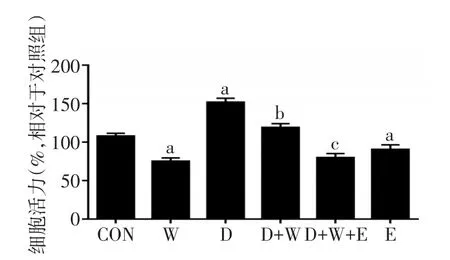
Fig.1 Changes of cell viability detected by MTT assay in six groups图1 MTT法分析各组细胞活力变化
2.2 杜拉鲁肽改善wortmannin诱导的细胞内Tau蛋白的过度磷酸化 (1)wortamannin和杜拉鲁肽干预后对细胞内总Tau蛋白没有明显影响(F分别为0.333和0.253,均P>0.05),Ex9-39干预也没有明显影响(t=0.182,P>0.05)。(2)wortamannin干预后Tau蛋白pS262、pS396和pS404位点磷酸化表达明显升高(F分别为267.814、196.761、510.729,均P<0.01),而杜拉鲁肽干预后Tau蛋白上述3个位点的磷酸化表达下降(F分别为219.231、119.028、15.182,均P<0.01),杜拉鲁肽和wortamannin存在交互效应,D+W组Tau蛋白pS262、pS396和pS404位点磷酸化表达下降(F交互分别为 49.190、24.290、5.466,均 P<0.05)。Ex9-39干预后 Tau蛋白 pS262、pS396和pS404位点磷酸化表达明显升高(t分别为7.994、14.241、15.761,P<0.01),D+W+E组tau蛋白上述位点的磷酸化表达高于D+W组(t分别为4.899、5.765、2.440,均P<0.05),见图2。
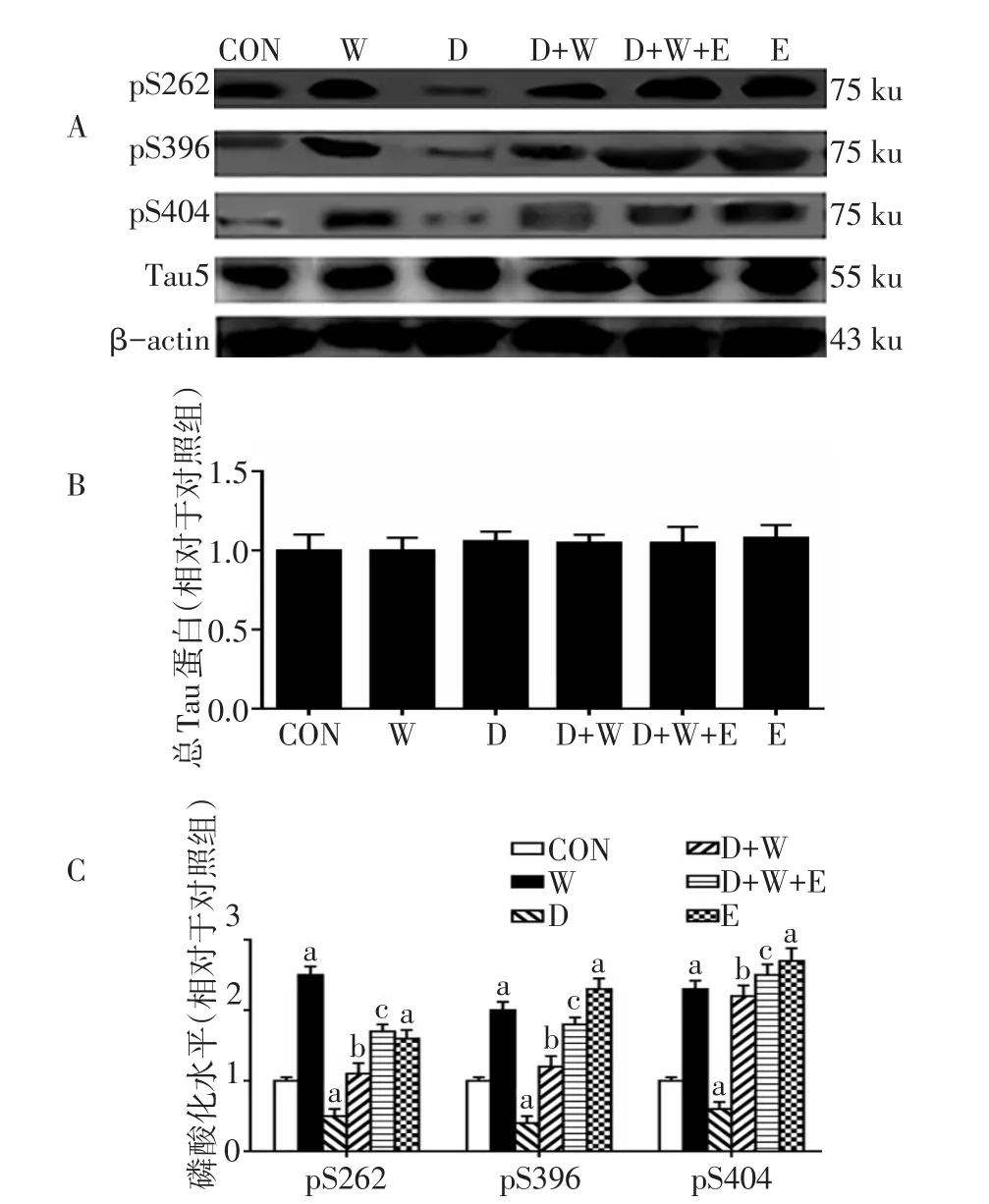
Fig.2 Expressions of total Tau protein and pS262,pS396 and pS404 detected by Western blot assay in SH-SY5Y cells图2 Western blot检测SH-SY5Y cells Tau蛋白及磷酸化表达
2.3 杜拉鲁肽减少wortmannin诱导的细胞内NF-H/M磷酸化水平 (1)wortamannin和杜拉鲁肽干预后对细胞内NF-H和NF-M神经丝均没有明显影响(F分别为0.300、2.700和0.370和0.333,均P>0.05),Ex9-39干预也没有明显影响(t分别为1.082和1.549,P>0.05)。(2)wortamannin干预后NF-M/H磷酸化水平上升(F分别为80.667、235.2,均P<0.01),而杜拉鲁肽干预后NF-M/H磷酸化水平下降(F分别为 112.667、388.8,均 P<0.01),杜 拉 鲁 肽 和wortamannin存在交互效应,D+W组NF-M/H磷酸化水平下降(F交互分别为43.200和16.667)。Ex9-39干预后NF-M/H磷酸化水平上升(t分别为6.991、6.662,P<0.05),D+W+E组NF-M/H磷酸化水平高于D+W组(t分别为10.568、10.067,均P<0.05)。见图3。
2.4 杜拉鲁肽提高p-PI3K(p85)和p-GSK3β(S9)的表达水平 (1)wortamannin和杜拉鲁肽干预后对细胞内胰岛素信号通路相关蛋白PI3K均没有明显影响(F分别为0.110和0.143,均P>0.05),Ex9-39干预也没有明显影响(t=1.820,P>0.05)。对GSK3β均没有明显影响(F分别为3.900和0.208,均P>0.05),Ex9-39干预也没有明显影响(t=0.643,P>0.05)。(2)wortamannin干预后p-PI3K和p-GSK3β水平下降(F分别为274.766、49.693,均P<0.01),而杜拉鲁肽干预后p-PI3K和p-GSK3β水平上升(F分别为 140.187、30.061,均 P<0.01),杜 拉 鲁 肽 和wortamannin存在交互效应,D+W组p-PI3K和p-GSK3β水平上升(F交互分别为14.020和13.800,均P<0.05)。Ex9-39干预后p-PI3K和p-GSK3β降(t分别为3.326和5.140,均P<0.05),D+W+E组NFM/H磷酸化水平低于D+W组(t分别为5.410和2.970,均P<0.05)。见图4。
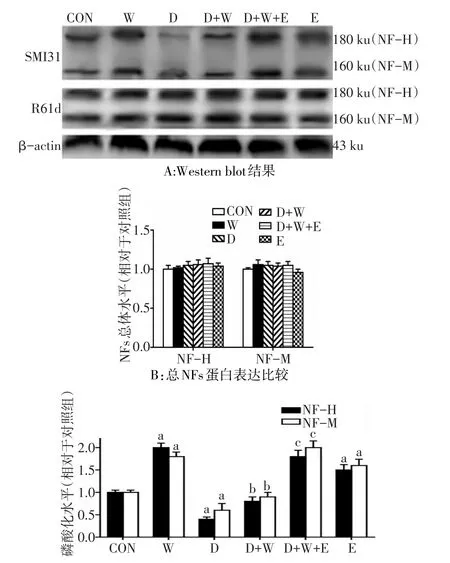
Fig.3 The phosphorylation levels of NF-H and NF-M detected by Western blot assay in SH-SY5Y cells图3 Western blot检测SH-SY5Y细胞NF-H/M蛋白及其磷酸化水平
3 讨论
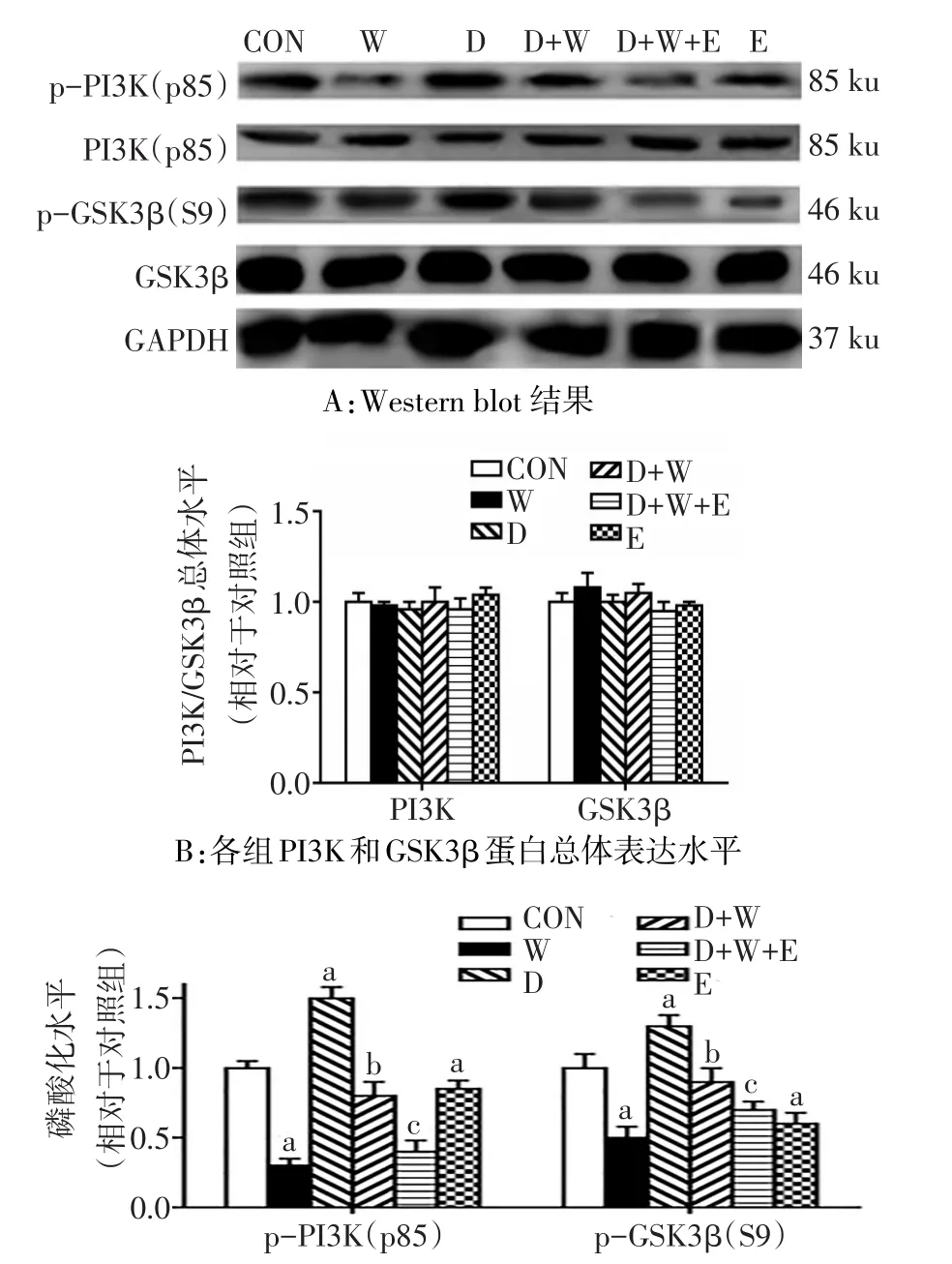
Fig.4 The phosphorylation levels of PI3K/GSK3β signaling pathways detected by Western blot assay in SH-SY5Y cells图4 Western blot检测SH-SY5Y细胞PI3K/GSK3β的磷酸化水平
AD是一种慢性神经退行性疾病并最终导致患者的痴呆和死亡,伴随着老龄化的加剧和慢性疾病如糖尿病和高血压发病率增加,我国AD绝对患病人数呈急剧上升趋势,严重影响老龄人群的健康和生存质量,因此预防和治疗AD具有重要意义[13]。然而95%的AD病例属于散发型,多种因素如遗传易感性、代谢改变和环境因素等可能导致散发性AD的发生,尽管在AD的发病机制研究方面已取得一些进展,特别是发现Aβ过度产生和聚集可导致AD样病理改变,然而没有在此基础上开发出有效的治疗药物[14-15]。本研究所用的SH-SY5Y细胞是人神经母细胞瘤SK-N-SH的亚型,与神经元有某些相似的生理功能,是一种良好的细胞模型,广泛应用于神经退行性变的疾病研究中[16]。本实验通过给予SHSY5Y细胞PI3K抑制剂wortmannin建立AD样细胞模型,可以明显地观察到过度磷酸化细胞骨架蛋白Tau和NFs在细胞内的聚集和细胞的形态学改变等类似AD退行性变的病理特点,以此为模型可以观察治疗糖尿病新药杜拉鲁肽的神经保护作用及机制,阐明其与AD样病理改变及PI3K信号通路的关系,为进一步在动物实验中应用提供理论依据[17]。
GLP-1作为一种肠促胰岛素,其相关药物如利拉鲁肽和艾塞那肽等用于治疗2型糖尿病,受到广泛关注[18-19]。GLP-1R在脑内广泛存在,在食欲抑制、神经元活性的调节、海马神经元回路的活性调节等过程中有重要作用[20-22]。GLP-1在中枢神经系统中起着促进神经元生长和修复,减少炎症、凋亡和氧化应激的神经营养因子的作用[23-24]。但是GLP-1在体内很容易被二肽基肽酶Ⅳ(dipeptidylproteaseⅣ,DPP-4)降解,GLP-1类似物杜拉鲁肽的氨基酸序列和分子质量有所改变,半衰期延长至48 h,每周注射1次,可促进胰岛素分泌,降低血糖,保护胰岛β细胞[25-26]。脑内 GLP-1通过和 GLP-1R 结合激活GLP-1信号,与神经元的生长、存活,血糖的调控及学习记忆功能密切相关[27]。GLP-1R基因缺失的小鼠学习能力下降、脑组织糖代谢减弱并伴有神经细胞的抗损伤能力下降,而海马过表达GLP-1R以及外源给予GLP-1类似物[Ser(2)]exendin(1-9)可改善小鼠的学习记忆能力、调节糖代谢和保护神经的凋亡,提示GLP-1信号在学习记忆和葡萄糖稳态(glucose homeostasis)调节中起关键作用[28]。GLP-1类似物利拉鲁肽虽然半衰期为13 h,但需每天给药,这就限制了GLP-1的脑内应用。杜拉鲁肽对神经细胞的作用鲜见报道[29-30]。本研究显示,与对照组和wortmannin组相比,杜拉鲁肽能提高神经细胞的活力,上调p-PI3K(p85)和p-GSK3β(S9)信号通路磷酸化水平,并下调了Tau蛋白pS262、pS396和pS404位点的磷酸化水平和NF-H/M磷酸化水平,表明杜拉鲁肽可通过与GLP-1R结合激活PI3K信号通路相关分子,改善胰岛素信号通路,增加细胞的糖代谢,影响Tau和神经丝蛋白的磷酸化和聚集,对抗神经细胞的退行性变,深入探讨其作用和机制可为AD防治药物的研究提供新的思路。
