高节竹和刺黑竹的光合特性分析
2018-07-18李欣欣陈凌艳陈礼光荣俊冬何天友郑郁善
李欣欣,陈凌艳,陈礼光,荣俊冬,何天友,郑郁善
(1.福建农林大学林学院,福建福州 350002; 2.福建农林大学园林学院,福建福州 350002)
近年来,随着城市建设的不断增强,城市内部的钢筋水泥陆续增多,绿色植物建设无疑成为一项必要的工程。竹类植物以其独具特色的观赏价值、文化内涵等优势在城市绿化和园林建设等方面逐渐受到重视,成为当今园林绿化植物的新宠儿,在艺术环境塑造中占有重要的地位[1]。伴随着竹类装饰地位的日益重要,观赏竹的研究也不断发展,国内外学者也纷纷研究了常见的青竹、淡竹、鼓节竹等观赏竹[2-4]。洪茜以园林中常见的18种观赏竹为对象,从观赏竹的绿量、滞尘效应、固碳释氧效应以及净化大气化学性污染物效应4个方面,对观赏竹净化环境功能进行定量测定与分析研究,筛选出了净化环境效果较好的观赏竹种[5];王啸晨等对白纹阴阳竹和白纹椎谷笹2种观赏竹的竹叶进行了形态结构观察和分析,发现其叶绿体中有大量的淀粉粒,可能具备抗低温能力[6]。光合作用是植物与外界进行交流的重要通道,是植物生存发展的基础[7]。谭向峰等对黄栌和小叶白蜡的光合特征进行了研究,发现黄栌对强光的适应性要高于小叶白蜡[8];韩拓等运用便携式光合仪对西北干旱地区的绿洲经济作物葡萄树叶进行了季节性光合作用分析,结果发现,夏季的净光合速率随着光照度的增加呈现先上升后平稳的趋势[9];蔡金桓等对狗牙花、红背桂和花叶假连翘在盐胁迫下的光合反应作了测定,最后得出,红背桂在盐胁迫下仍能维持较高的光合特性,适合在盐碱地中栽培,而花叶假连翘在盐胁迫下的光合能力弱,不适于在盐碱地中栽培[10]。针对竹类的光合特性研究也比较多[11-17]。高节竹(Phyllostachysprominens)和刺黑竹(Chimonobambusa.purpure)原产地都非福建地区,适当的引种可以增加本地观赏竹种类,丰富该地观赏竹品种。夏季是光合作用反应最为强烈的季节,本试验通过对7月高节竹和刺黑竹光合特性的测定,旨在为2竹种的园林绿化、适宜的生存环境和高效引种栽培提供理论依据。
1 材料与方法
1.1 研究区概况
试验材料取于福建农林大学中华名特优植物园内(简称中华园)。福建农林大学位于福建省福州市(地理位置25°15′~26°39′N,118°08′~120°31′E),属于亚热带季风气候,气候温暖湿润,四季常青,无雪,霜冻少,日照时数长,降水充沛,年均温为 16~20 ℃,相对湿度约达77%,风向以东北风为主,夏季以偏南风为主。中华园约占地7 hm2,地势平坦,本试验的竹种都是同一时期移植到中华园内的,所有的气压、海拔、土壤等自然环境条件均一致。本试验竹种为刚竹属(Phyllostachys)的高节竹(P.prominens)和寒竹属(Chimonobambusa)的刺黑竹(C.purpure)。
1.2 研究方法
选择在2011年7月无风少云的晴朗天气进行光合日变化测定[18-20],选取长势较好的植物各3株,每株的年龄、胸径与树高等基本一致,采用美国LI-COR公司生产的Li-6400便携式植物光合测量仪测定净光合速率、蒸腾速率、气孔导度、胞间CO2浓度、表观量子效率等,每次测量时选取5个数值,最后取其平均值。测定时的光照度梯度依次为0、10、30、50、70、100、200、300、500、800、1 000、1 200、1 800 μmol/(m2·s),测定从高光照度依次递减,测定前使试验样品在光照下诱导 20 min,测定时的CO2浓度设置为(380±20)μmol/mol,当其变化在0.5 μmol/mol范围内时连续记录5个值。
1.3 数据分析处理
运用Excel和SPSS 19.0对数据进行初步整理和分析,然后依据植物光合作用对光响应的指数方程Pn=Pn,max(1-C0·e-αI/Pn,max)拟合竹子的光响应曲线,其中,C0为度量弱光下净光合速率趋于0的指标,即C0=1-(P0/α);Pn,max为最大净光合速率;α为植物光合作用对光响应曲线在I=0时的斜率,即光响应曲线的初始斜率,也称为初始量子效率;e为自然数,其值可取为2.72;I为光照度[21-22]。
光合水分利用效率(WUE)根据Pn/Tr来计算;光补偿点(LCP)为净光合速率为0时的光照度,假定光饱和点(LSP)为最大净光合速率的99%,计算公式为LCP=Pn,max·lnC0/α,LSP=Pn,max·ln(100C0)/α[23]。
2 结果与分析
2.1 净光合速率对光照度的响应
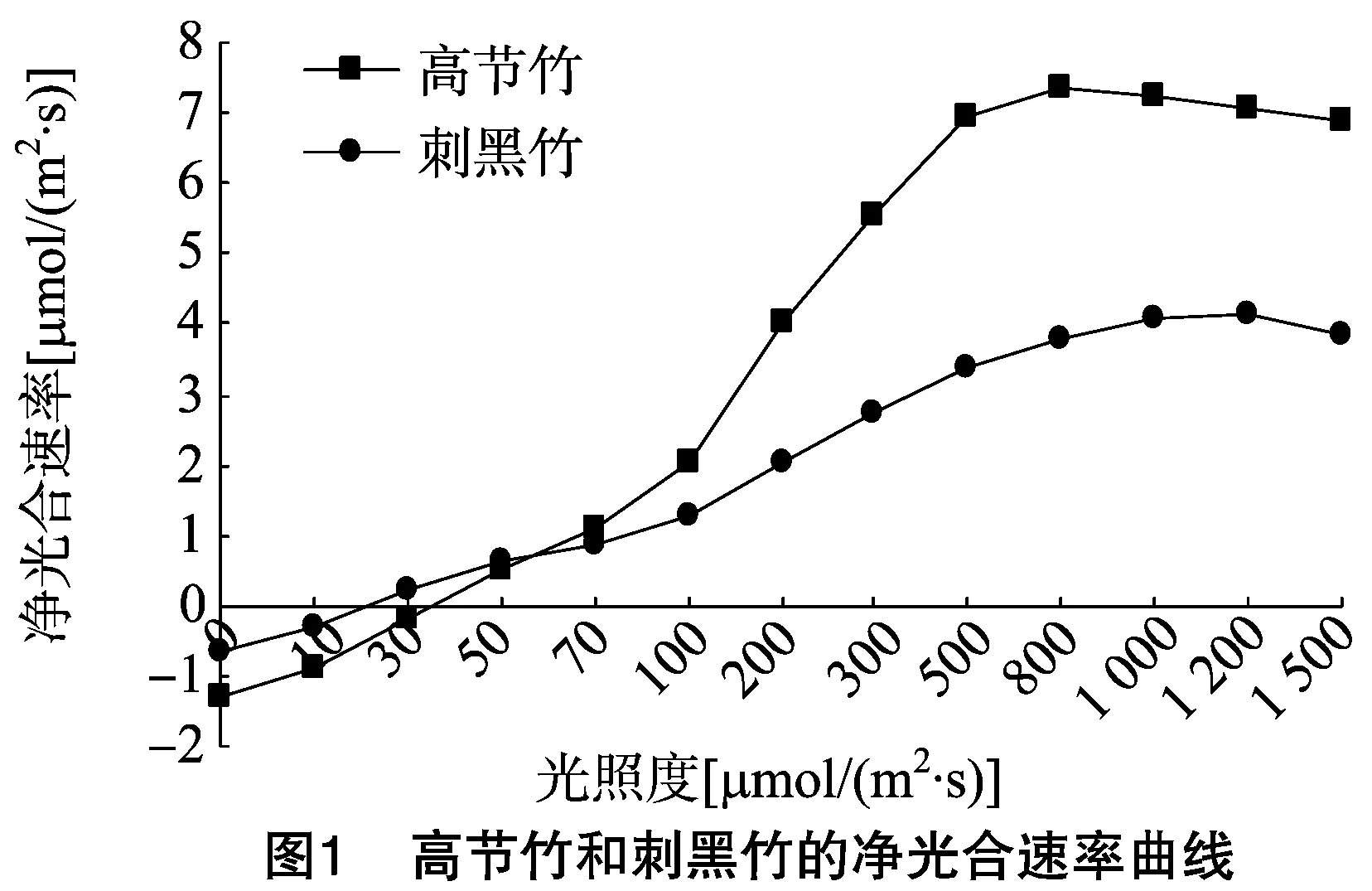
如图1所示,高节竹和刺黑竹的净光合速率都是随着光照度的增加而不断上升,在到达一定峰值后趋于平稳。当光照度为0~100 μmol/(m2·s)时,高节竹和刺黑竹的净光合速率都随着光照度的增加而缓慢上升;当光照度为100~800 μmol/(m2·s) 时,高节竹的净光合速率随着光照度的增加急速上升,在800 μmol/(m2·s)时达到峰值。而刺黑竹在光照度100~1 200 μmol/(m2·s)时净光合速率随光照度的增加急速上升,在光照度为1 200 μmol/(m2·s)时达到最大值。高节竹和刺黑竹的光合速率差异较大,虽然两者都是随着光照度的增加而增加的,到达各自的峰值后趋于平稳然后逐渐下降,但高节竹的净光合速率明显高于刺黑竹,高节竹的净光合速率的最大值为7.36 μmol/(m2·s),刺黑竹的最大净光合速率为4.13 μmol/(m2·s),高节竹的最大净光合速率是刺黑竹最大净光合速率的1.8倍左右。
2.2 蒸腾速率对光照度的响应
如图2所示,高节竹的蒸腾速率随着光照度的增加缓慢上升,然后逐渐趋于平稳,刺黑竹在光照度0~50 μmol/(m2·s) 时,蒸腾速率随着光照度的增加而缓慢下降,从 50 μmol/(m2·s) 开始,随着光照度的增加呈缓慢上升趋势,在光照度为1 600 μmol/(m2·s)时有急速上升趋势。高节竹和刺黑竹都是在光照度为1 800 μmol/(m2·s)时达到最大值,此时高节竹的蒸腾速率为2.91 mmol/(m2·s),刺黑竹的蒸腾速率为1.76 mmol/(m2·s),高节竹的蒸腾速率比刺黑竹的蒸腾速率大。
2.3 气孔导度对光照度的响应
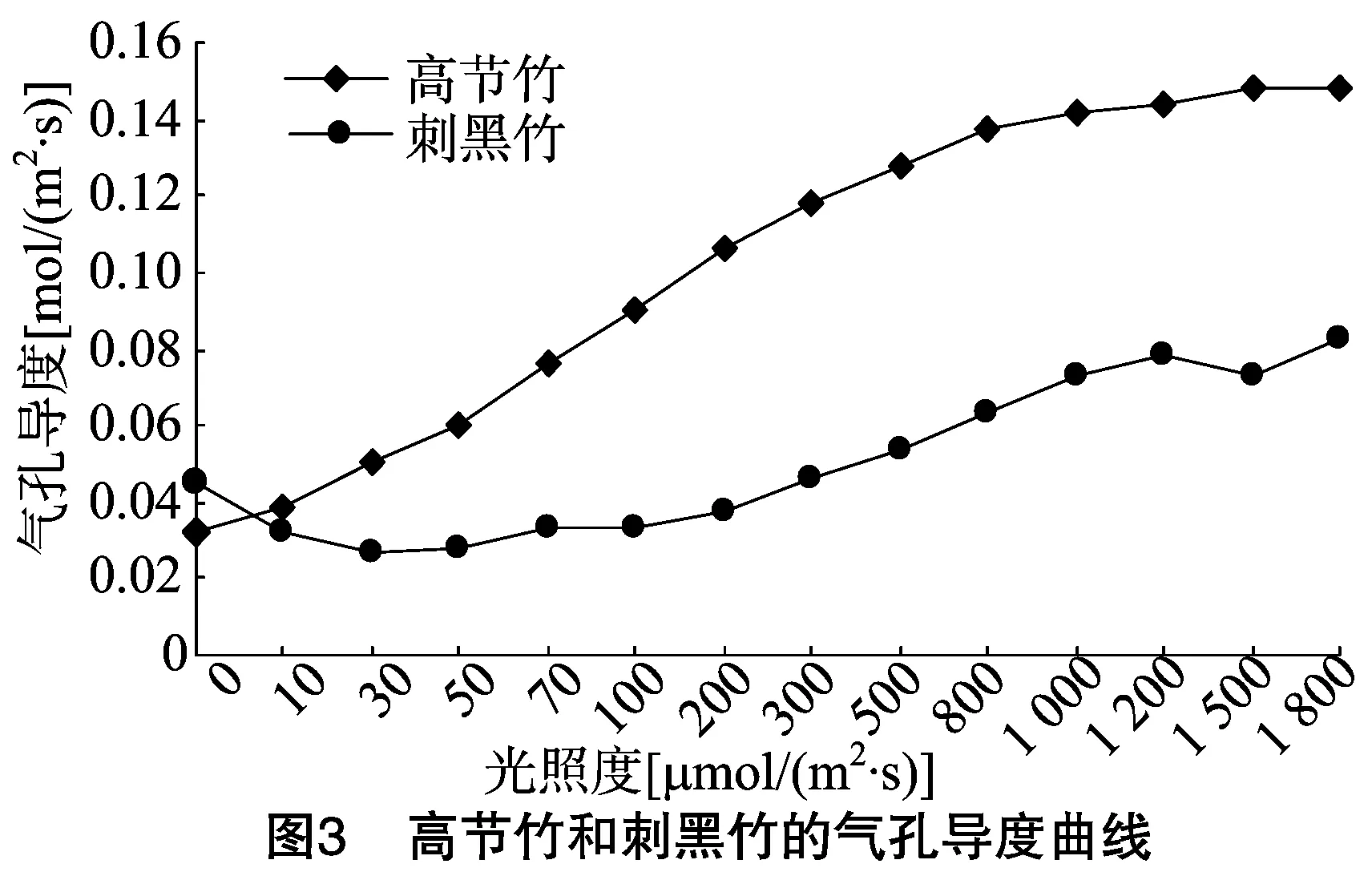
气孔导度是植物的气孔对水蒸气和CO2的传导能力,表示植物气孔的张开程度,影响植物进行光合作用、呼吸作用和蒸腾作用。气孔导度愈大,表示气孔张开得愈大,水蒸气和CO2就可以顺利地通过气孔进行传输。因而植物可以通过改变气孔导度的大小来控制植物内部与外界水蒸气和CO2的交换,从而调节植物的光合速率和蒸腾速率[24]。由图3可知,高节竹的气孔导度随着光照度的增加而迅速上升,然后趋于缓慢增长。刺黑竹的气孔导度与高节竹相比较为复杂,在光照度为0~40 μmol/(m2·s)时,气孔导度呈下降趋势,之后随着光照度的增加缓慢上升,在光照度为1 200 μmol/(m2·s) 时又下降,在光照度为1 200~1 500 μmol/(m2·s)范围是下降的,从1 500 μmol/(m2·s)开始再次上升。高节竹的气孔导度高于刺黑竹,这也解释了高节竹的净光合速率和蒸腾速率都高于刺黑竹的原因,说明植物的气孔导度与植物的净光合速率、蒸腾速率具有密切关系,有很强的相关性和一致性。

2.4 胞间CO2浓度对光照度的响应
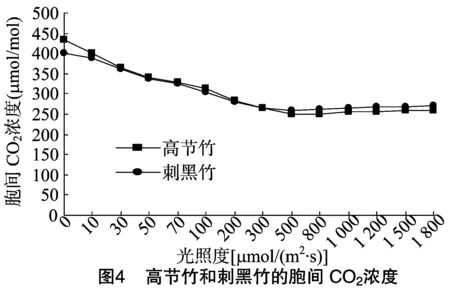
细胞间CO2的浓度,是外界CO2进入叶肉细胞过程中所受各种驱动力和阻力以及叶片内部光合作用和呼吸作用的最终平衡结果[25]。胞间CO2浓度与植物叶片周围空气的阻力、气孔导度、叶肉导度和叶肉细胞的光合活性有关,当空气中的CO2浓度增高时,导致胞间CO2的浓度升高;当空气中的CO2浓度降低时,胞间CO2的浓度也随着降低。由图4可知,当光照度在0~300 μmol/(m2·s)时,随着光照度的增加,胞间CO2浓度逐渐降低,光照度从300 μmol/(m2·s)开始,胞间CO2浓度逐渐趋于稳定,变化幅度较小。高节竹和刺黑竹两者的变化趋势接近,只是在光照度为0时高节竹的胞间CO2浓度大于刺黑竹。
2.5 水分利用率对光照度的响应
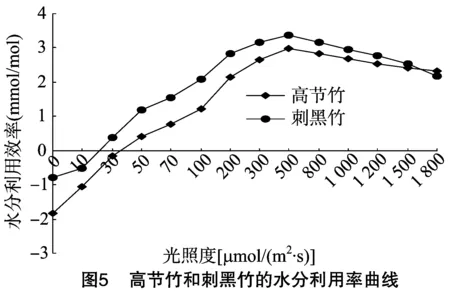
由图5可知,当光照度在50 μmol/(m2·s)以下时,高节竹的水分利用率为负值,光照度在30 μmol/(m2·s)以下时,刺黑竹的水分利用率为负值。随着光照度的增加,高节竹和刺黑竹的水分利用率都随之起伏上升,在光照度约为 600 μmol/(m2·s)时两者的水分利用率都逐渐平缓下降,即两者都有先增后减的趋势。从图5可以看出,高节竹的水分利用率低于刺黑竹。
2.6 竹种的光合作用参数
由表1可以看出,高节竹的表观量子速率为 0.032 9 μmol/(m2·s),刺黑竹的为0.019 0 μmol/(m2·s),高节竹的表观量子速率大于刺黑竹。高节竹的光补偿点为 35.85 μmol/(m2·s),刺黑竹的为30.64 μmol/(m2·s),高节竹的光补偿点比刺黑竹的高约5 μmol/(m2·s)。高节竹的光饱和点为 1 066.06 μmol/(m2·s),刺黑竹的光饱和点为 1 031.66 μmol/(m2·s),高节竹的光饱和点比刺黑竹的高约34 μmol/(m2·s)。综合来看,高节竹的表观量子速率、光补偿点、光饱和点都是高于刺黑竹的,说明高节竹具有适应强光的能力。

表1 高节竹和刺黑竹的光合作用参数
3 结论与讨论
光照度对植物光合作用、形态结构、生理机制等都有较大的影响。了解植物内部的各个生理指标对于促进植物生长具有重大意义。掌握影响植物进行光合作用的因子如净光合速率、蒸腾速率、气孔导度、胞间CO2浓度和水分利用率,进而分析出植物的表观量子速率、光补偿点和光饱和点,根据这些重要分析结果来为植物寻得合适的生长环境以及促进其生产力达到最大值。
高节竹的净光合速率、蒸腾速率和气孔导度都高于刺黑竹,胞间CO2浓度两者几乎一致,刺黑竹的水分利用效率高于高节竹。植物的气孔导度和蒸腾速率都与植物的净光合速率呈正相关关系,都是随着光照度的增加逐步上升,到达其峰值后平稳下降。植物蒸腾速率越大,表明植物与外界交换气体和水分功能越发达,降低植物叶片的温度,使植物在接受强光时不至于受伤害。影响植物蒸腾速率的主要因素就是植物叶片的气孔导度,叶片气孔的大小影响植物蒸腾速率的快慢,气孔导度就是叶片气孔对外界水蒸气、CO2等气体的传导度。蒸腾速率和气孔导度共同影响植物光合速率的快慢。高节竹的蒸腾速率和气孔导度都大于刺黑竹的,进而光合速率也比刺黑竹快,三者相互影响,任何一个因素的变化都将会影响其他因素的变化。
比较2个竹种的水分利用率可以发现,刺黑竹的水分利用率高于高节竹的,水分利用率的高低说明了植物对水分的利用情况,水分利用率高代表节约水资源,刺黑竹的水分利用率高,表明刺黑竹对水分的要求相比高节竹低,可以种植在相对少水的地方。植物的表观量子效率表示植物对弱光的利用能力,光补偿点表示植物对弱光的适应能力,光饱和点表示植物对光的喜好程度[23]。由本研究结果可知,高节竹的表观量子率、光补偿点和光饱和点都高于刺黑竹,水分利用率低于刺黑竹,所以高节竹适于生长在水热条件较好的地区,对水分和光的要求比较高,适宜种植在山的阳坡和光照水分条件较好的园区内。刺黑竹对光照和水分的相对要求较低,可以种植在山坡北麓和相对背阴的园区。分析竹种的生理特性,了解各竹种对光照和水分等外界环境条件的要求,选择适宜的生长条件予以种植,有助于充分发挥该竹种的观赏价值和生产价值。
