梨自交花粉原位萌发观察及不亲和性强度研究
2018-07-18金子明王国明柯亚琪石苏利
金子明,王国明,柯亚琪,石苏利,吴 磊,谷 超
(南京农业大学梨工程技术研究中心,江苏南京 210095)
蔷薇科梨属(PyrusL.)植物普遍存在由S基因位点控制的配子体型自交不亲和(gametophytic self-incompatibility)现象,大多数品种表现为自花授粉不亲和以及低结实率的特征[1]。大量研究表明,梨自花授粉不亲和性主要是由花柱分泌的S糖蛋白引起的,这种S糖蛋白能够分解自花授粉花粉管的RNA,导致花粉管停止生长并进入程序性死亡[2-3]。也有人认为该反应由S位点(S-locus)的1对S等位基因控制,即由雌蕊和花粉的S基因控制,分别为S-RNase、S-locusF-box/S-haplotype-specificF-box基因[4]。当花粉的S基因与雌蕊的1对S等位基因之一的S基因相同时,自花花粉能够在柱头上萌发,但花粉管沿花柱向子房伸长过程中会被花柱内的S糖蛋白抑制,而不能完成受精坐果[5],这种普遍存在的机制能够抑制品种自交,促进异花授粉。在各种自然因素的作用下,自交不亲和植物也会发生自交亲和性突变,一种是花柱突变,如日本梨品种奥嗄二十世纪是二十世纪(Nijisseiki,S基因型S2S4)的突变体,其中1个花柱S基因,即S42RNase基因发生变异,导致奥嗄二十世纪的花柱不能识别自己的花粉[6]。另一种自交亲和性突变就是花粉发生突变,即花粉失去自交不亲和(self-incompatibility,简称SI)识别功能,而形成花粉SI突变体[5]。如吴华清等研究的金坠梨是鸭梨的芽变品种,金坠梨自花授粉结实率高达76%,自花的花粉管能正常生长至花柱基部,由于花粉S基因发生突变,导致自交不亲和性功能的丧失,表现出自花授粉能够结实[7]。梨属于配子体型自交不亲和果树,绝大多数品种自花授粉结实率极低,在生产上必须配置授粉树以及花期人工辅助授粉,才能完成授粉受精,获得经济产量,但是同时也增加了生产成本[4]。尽管如此,也常因授粉树配置不合理、人工授粉不及时或是花期不良的气候条件等问题影响授粉受精而造成减产[5],而选育自花结实性品种是解决这些问题的重要途径之一。加深对梨自交不亲和性的研究,不仅有助于更好地了解这一生命现象,还可以为果树育种和生产提供可靠的理论依据。同时为进一步阐明梨自交亲和性突变机制,深入研究自花花粉管被抑制的机制,从形态学角度探明花粉管在花柱中的生长特性,以及不同品种自交不亲和强度分析统计,为显花植物防止自交衰退这一遗传机制提供一些试验统计数据。因此,对不同品种的梨树自交不亲和性强度的研究,不仅具有理论价值,而且有潜在的实践意义。
1 材料与方法
1.1 材料准备
本试验于2014年3月至2016年6月,分别于江苏省农业科学院梨资源圃和南京农业大学江浦农场园艺试验站梨资源圃内进行,共有256个品种进行套袋处理。每个品种随机选取6朵发育正常的并且处于大蕾期的花朵进行硫酸纸套袋处理。待花开后花药散粉时,轻轻摇动纸袋或树枝,使其自交授粉。为了确保每朵花自交授粉成功,从套袋后第2天起,每天摇动纸袋或树枝2次。5 d后采取套袋花朵,从基部截取花柱并迅速置于FAA(甲醛 、冰乙酸、70%乙醇体积比 5 ∶5 ∶90)固定液中,4 ℃保存。
1.2 花粉管原位荧光显微观察并拍照
花粉原位萌发的荧光显微观察技术参照吴巨友等的方法[8],并稍有改动:从FAA固定液中取出样品,用蒸馏水多次清洗后,放入2 mol/L NaOH溶液中软化,65 ℃水浴40 min,使组织软化;再次清洗多次后,用0.1%苯胺蓝水溶液(用0.1% K3PO4溶液配制)避光室温染色4~6 h。多次清洗后,滴甘油压片,用宏观变倍体式荧光显微镜(型号:MVX10,日本OLYMPUS公司)观察并拍照。
1.3 数据处理
利用Image-ProPlus6.0软件测量花柱中花粉管的生长长度及其花柱的长度,利用Excel 2007统计软件计算不同品种花粉管伸长长度占花柱全长的比值,并进行统计分析。
1.4 坐果率调查
于2016年选取筛选后的梨品种进行自花授粉坐果率调查。在开花前1周,对发育一致的花序进行疏花,只留 2~3个生长较好的花蕾,进行套袋处理。盛花期轻轻摇动套袋的花序,进行自花授粉,每天摇动纸袋或树枝2次。分别在盛花后20、30 d统计坐果率,坐果率大于20%被认为是自交亲和[9]。
2 结果与分析
2.1 花粉管生长特性观察
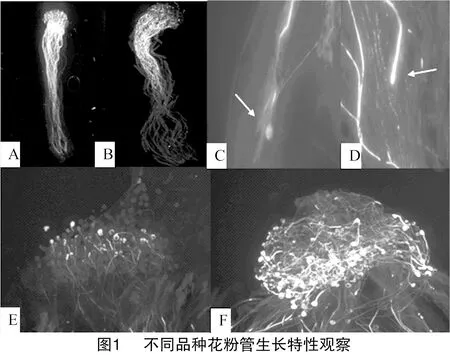
对梨不同品种花粉管原位荧光拍照观察发现,花柱中花粉管分布及数量表现为上部较多、中部较少、下部最少的梯度分布(图1-A、图1-B)。部分自花授粉的花粉管停止生长时会出现少量的尖端膨大和弯曲等变形现象,与陈迪新等研究观察结果一致[10](图1-C、图1-D)。同时发现自交不亲和性较强的品种花粉管虽然有少量穿过柱头,但不能进一步在花柱内生长,表现为花粉管在柱头上为畸形状、扭曲变形、先端膨大、杂乱无章等不亲和性现象(图1-E、图1-F)。
2.2 梨自交不亲和性强度
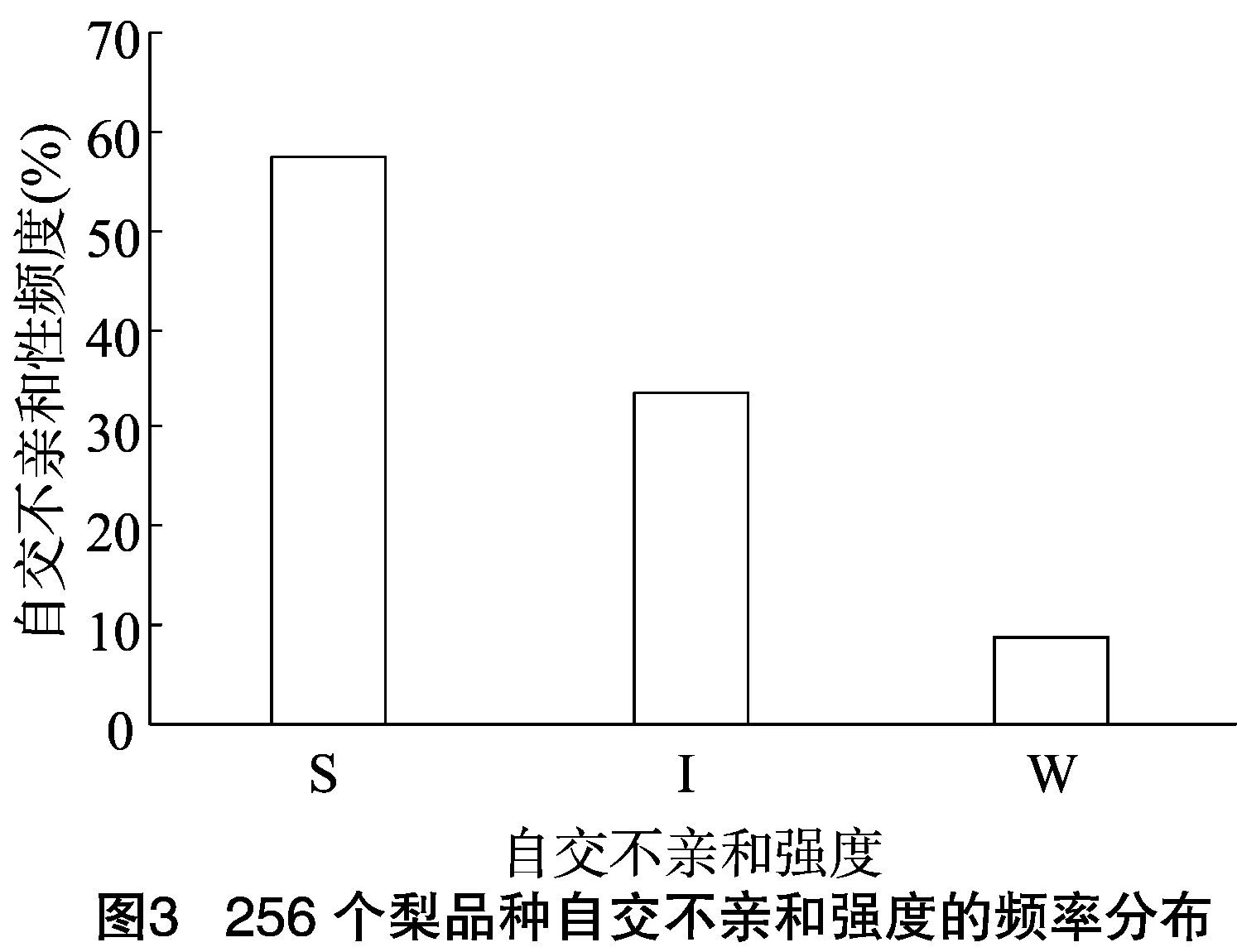
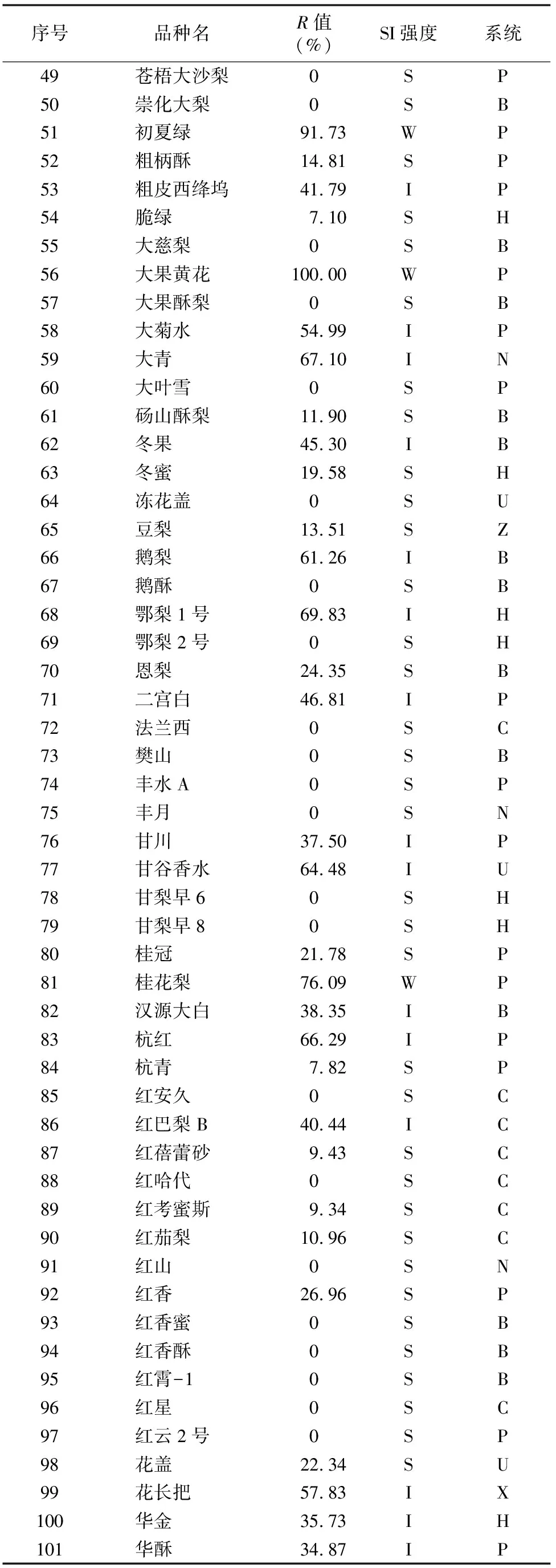
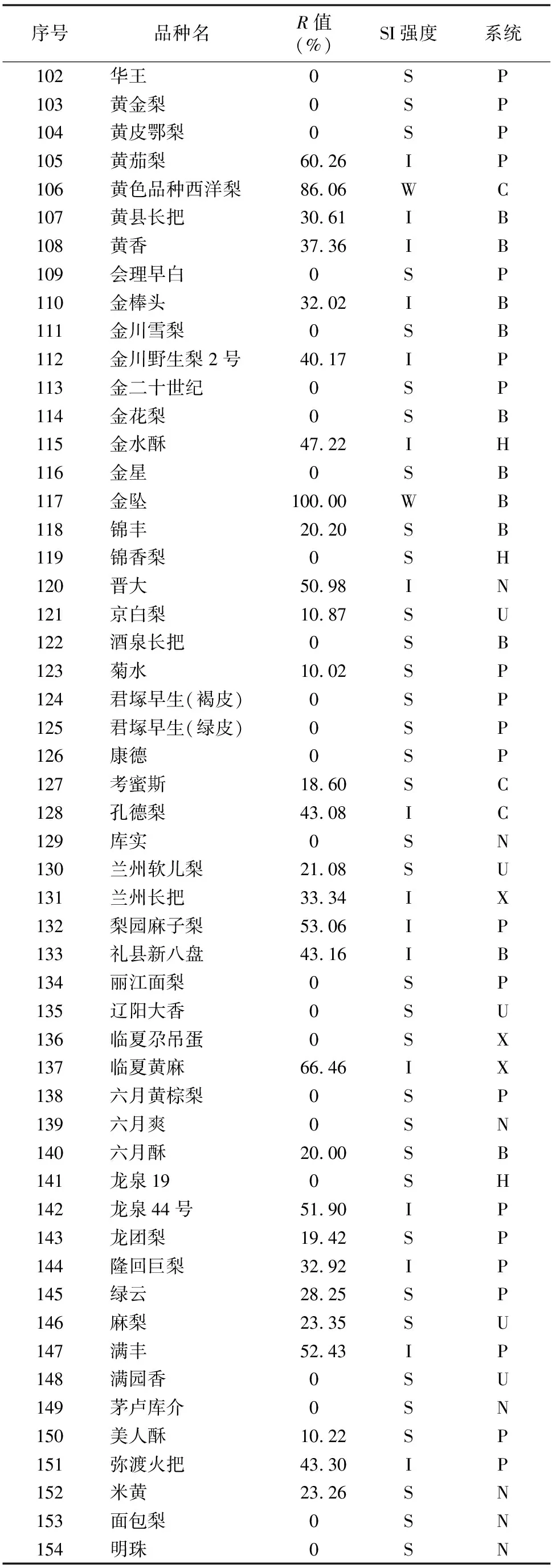
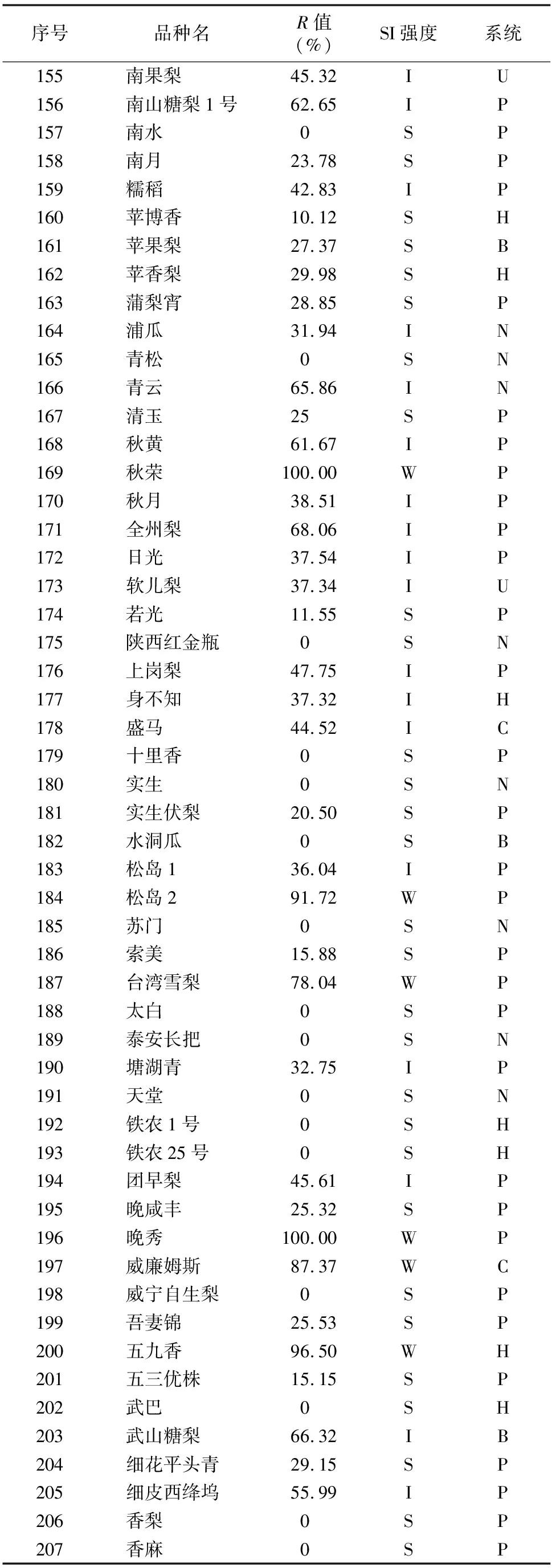
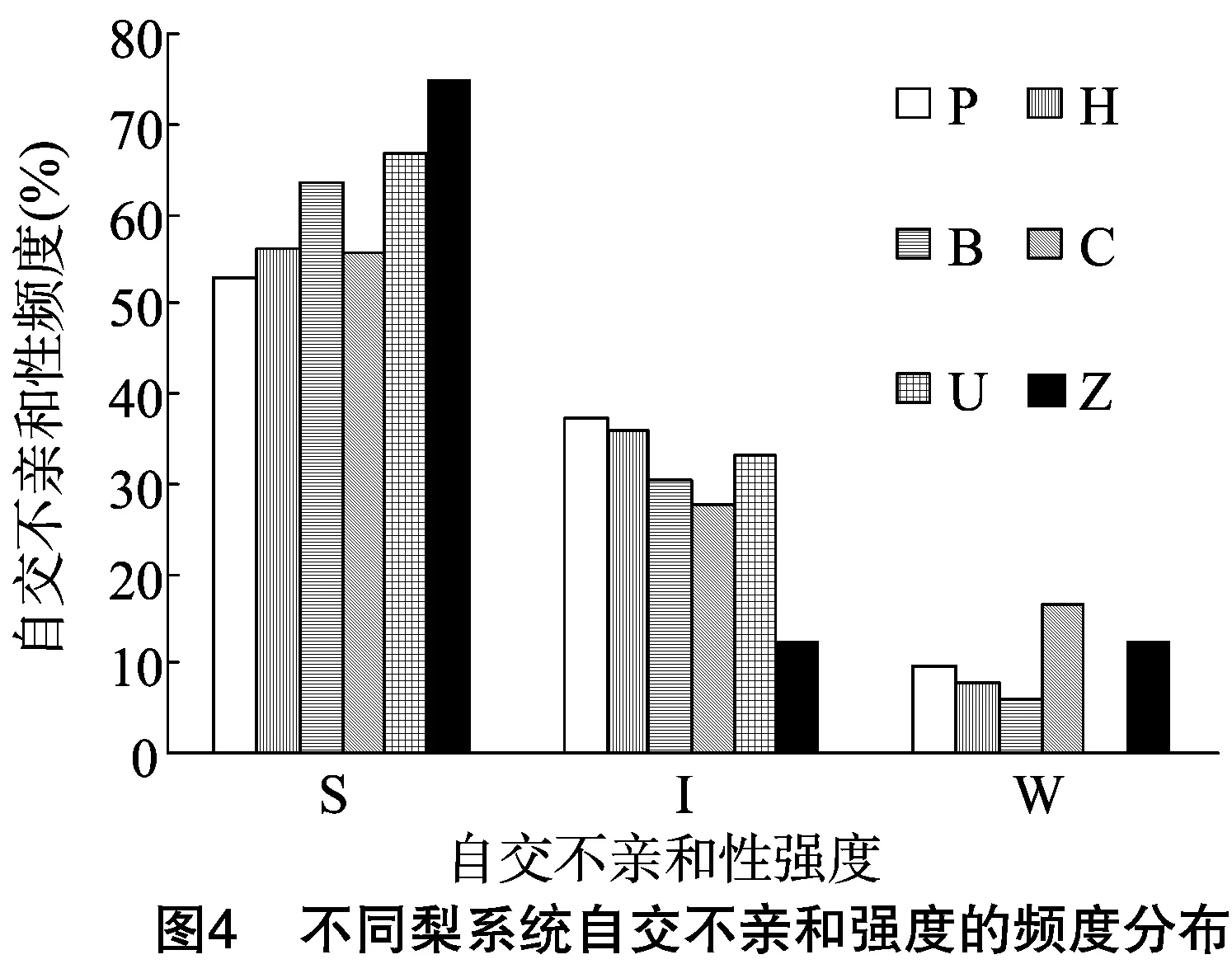
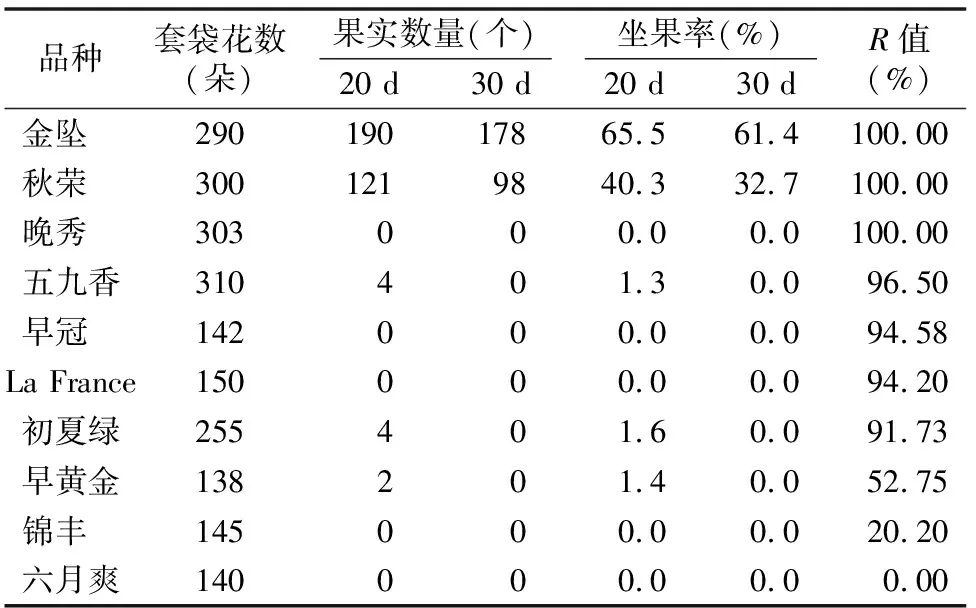
由于不同果树树种自交亲和与不亲和性的判定标准不同,且对于同一树种不同研究者掌握的尺度也不同。张绍铃等通过S糖蛋白含量高低来判断自交不亲和强度[11]。还有研究表明,自花授粉的花粉管在花柱中生长长度与其S糖蛋白含量显著相关[12]。本研究根据自交授粉花柱中花粉管生长长度占整个花柱的比例定义品种的自交不亲和强度R值,通过对R值进行分类,将梨供试品种的自交不亲和强度分为强(强,0≤R≤30%)、中(中,30% 由于梨自交不亲和的花粉能在柱头上萌发并进入花柱,但在花柱中的生长受到抑制,不能完成受精,本试验对套袋处理的256个梨品种从基部截取花柱,染色拍照并进行统计分析。结果显示,不同梨品种自交不亲和性强度差异明显,不亲和性强度变幅为0~100%,自交不亲和性强度R值强、中、弱参差不齐。花粉管可以完全长到基部,共发现6个极可能为自交亲和性的品种,只占调查总数的2.34%。本研究发现大多数品种花粉管不能长到基部(表1),符合梨属于自交不亲树种的基本属性,说明梨属于典型的自交不亲和性植物。这6个不同梨品种的R值为100%,其中闫庄鸭梨[13]、秋荣[14]、54S-135[14]、金坠梨[7]、大果黄花梨[15]是已报道的自交亲和性品种,而晚秀则是本试验通过自花花粉管发现的极可能为自交亲和性品种。这些品种的花粉管能在自交花柱中生长并长至花柱底部,至于能否完成下一步受精及坐果过程,需对这些品种进行进一步自花套袋座果率调查。通过鉴定自交不亲和强度R值,对后期自交亲和性品种进行了初步筛选。并且为后期探索花粉管生长长度与自交不亲和强度的深入研究提供了充足的基础数据。 对256个品种分析发现,其中自交不亲和性强度为强的有147个,占所调查总数的57.4%;自交不亲和性强度为中的有86个,占33.6%;自交不亲和性强度为弱的有23个,占9.0%。从频度分布角度分析,梨不同品种自交不亲和强度主要趋于强与中,花粉管能长到花柱基部的占少数,进一步验证了梨属于配子体自交不亲和植物(图3)。对256个品种所属系统归类和对不同系统自交不亲和性强度的分布频度进行统计比较,发现每个系统自交不亲和性强度R值的强、中和弱分布符合所调查的整体规律,自交不亲和性强的占大多数,而自交不亲和性弱的仅占少数(表1、图4)。同时对比发现,西洋梨与砂梨系统自交不亲和性强度R值弱的分布频度较其他系统高(图4),为发现更多的自交亲和性品种提供了方向。 表1 不同梨品种自交花柱中花粉管的生长长度占花柱的比例(R值)、自交不亲和性强度分类及所属系统 续表1 序号品种名R值(%)SI强度系统49苍梧大沙梨0SP50崇化大梨0SB51初夏绿91.73WP52粗柄酥14.81SP53粗皮西绛坞41.79IP54脆绿7.10SH55大慈梨0SB56大果黄花100.00WP57大果酥梨0SB58大菊水54.99IP59大青67.10IN60大叶雪0SP61砀山酥梨11.90SB62冬果45.30IB63冬蜜19.58SH64冻花盖0SU65豆梨13.51SZ66鹅梨61.26IB67鹅酥0SB68鄂梨1号69.83IH69鄂梨2号0SH70恩梨24.35SB71二宫白46.81IP72法兰西0SC73樊山0SB74丰水A0SP75丰月0SN76甘川37.50IP77甘谷香水64.48IU78甘梨早60SH79甘梨早80SH80桂冠21.78SP81桂花梨76.09WP82汉源大白38.35IB83杭红66.29IP84杭青7.82SP85红安久0SC86红巴梨B40.44IC87红蓓蕾砂9.43SC88红哈代0SC89红考蜜斯9.34SC90红茄梨10.96SC91红山0SN92红香26.96SP93红香蜜0SB94红香酥0SB95红霄-10SB96红星0SC97红云2号0SP98花盖22.34SU99花长把57.83IX100华金35.73IH101华酥34.87IP 续表1 序号品种名R值(%)SI强度系统102华王0SP103黄金梨0SP104黄皮鄂梨0SP105黄茄梨60.26IP106黄色品种西洋梨86.06WC107黄县长把30.61IB108黄香37.36IB109会理早白0SP110金棒头32.02IB111金川雪梨0SB112金川野生梨2号40.17IP113金二十世纪0SP114金花梨0SB115金水酥47.22IH116金星0SB117金坠100.00WB118锦丰20.20SB119锦香梨0SH120晋大50.98IN121京白梨10.87SU122酒泉长把0SB123菊水10.02SP124君塚早生(褐皮)0SP125君塚早生(绿皮)0SP126康德0SP127考蜜斯18.60SC128孔德梨43.08IC129库实0SN130兰州软儿梨21.08SU131兰州长把33.34IX132梨园麻子梨53.06IP133礼县新八盘43.16IB134丽江面梨0SP135辽阳大香0SU136临夏尕吊蛋0SX137临夏黄麻66.46IX138六月黄棕梨0SP139六月爽0SN140六月酥20.00SB141龙泉190SH142龙泉44号51.90IP143龙团梨19.42SP144隆回巨梨32.92IP145绿云28.25SP146麻梨23.35SU147满丰52.43IP148满园香0SU149茅卢库介0SN150美人酥10.22SP151弥渡火把43.30IP152米黄23.26SN153面包梨0SN154明珠0SN 续表1 序号品种名R值(%)SI强度系统155南果梨45.32IU156南山糖梨1号62.65IP157南水0SP158南月23.78SP159糯稻42.83IP160苹博香10.12SH161苹果梨27.37SB162苹香梨29.98SH163蒲梨宵28.85SP164浦瓜31.94IN165青松0SN166青云65.86IN167清玉25SP168秋黄61.67IP169秋荣100.00WP170秋月38.51IP171全州梨68.06IP172日光37.54IP173软儿梨37.34IU174若光11.55SP175陕西红金瓶0SN176上岗梨47.75IP177身不知37.32IH178盛马44.52IC179十里香0SP180实生0SN181实生伏梨20.50SP182水洞瓜0SB183松岛136.04IP184松岛291.72WP185苏门0SN186索美15.88SP187台湾雪梨78.04WP188太白0SP189泰安长把0SN190塘湖青32.75IP191天堂0SN192铁农1号0SH193铁农25号0SH194团早梨45.61IP195晚咸丰25.32SP196晚秀100.00WP197威廉姆斯87.37WC198威宁自生梨0SP199吾妻锦25.53SP200五九香96.50WH201五三优株15.15SP202武巴0SH203武山糖梨66.32IB204细花平头青29.15SP205细皮西绛坞55.99IP206香梨0SP207香麻0SP 续表1 序号品种名R值(%)SI强度系统208湘菊0SP209新九梨22.30SN210新世纪73.45WP211新兴0SP212新秀46.00IP213新鸭梨39.94IB214兴矮二号55.92IP215兴山10号0SP216兴山15号26.06SP217兴山16号53.56IP218兴山34号21.56SP219兴山8号22.86SP220幸藏55.99IP221雄古冬梨37.31IP222秀黄59.08IP223雪英56.22IH224鸭梨28.01SB225雅青68.49IH226闫庄鸭梨100.00WB227义乌子梨0SP228意大利7号0SC229油酥21.36SP230玉露香0SH231圆黄26.01SP232云和柿扁梨8.24SP233云林55.02IP234云南海东40.80IN235早冠94.58WH236早黄金52.75IP237早金酥0SH238早绿43.39IH239早三花0SP240早酥蜜0SB241早香脆31.12IB242长把酥0SP243浙12-435.15IP244浙2113.61SP245蔗梨15.54SH246真鍮43.80IP247芝麻梨60.08IP248祗圆85.74WP249中矮3号15.32SZ250中翠7.93SH251中梨1号24.37SH252中梨3号18.81SH253重阳红0SN254猪嘴巴24.84SP255筑波4967.37IP256筑水34.50IP 注:S(strong)代表自交不亲和强度强的品种,I(intermediate)代表中强度的品种,W(weak)代表弱强度的品种,B代表白梨(P.brestschneideri),C代表西洋梨(P.communis),H代表种间杂交(interspecific hybridization),N(unknow species)代表未知种类,P代表砂梨(P.pyrifolia),U代表秋子梨(P.ussuriensis),X代表新疆梨(P.sinkiangensis),Z(stock species)代表砧木类型。 对于自交亲和性强度极弱的,笔者选择了R>90%的品种(金坠、秋荣、晚秀、五九香、早冠、初夏绿)进一步探索自花授粉结实率,结果发现,金坠、秋荣、晚秀3个品种R值为100.00%,其中以已报道的自交亲和品种金坠和秋荣作为对照。同时笔者也对自交不亲和强度为中的早黄金、自交不亲和强度为强的锦丰和六月爽多个品种进行了自花套袋试验。表2结果表明,金坠和秋荣在花后20、30 d坐果率都高于20%,而其他品种自花套袋的品种基本没有坐果。金坠和秋荣自花花粉管长到花柱最低端,并且坐果率大于20%,说明金坠和秋荣为自交亲和性品种,同时通过自交不亲和强度R值来初步筛选自交亲和性品种的可靠性得到了一定的验证。然而,并不是所有R值为100.00%的品种都是自交亲和性品种,本试验中,初步鉴定晚秀R值为100.00%,但通过自花套袋发现坐果率为0。其他自交亲和性强度为强与中的品种自花套袋不结实,或者结实率极低,为自交不亲和品种,进一步说明通过R值在海量的资源中初步筛选自交亲和性品种方法的优越性及可靠性。 表2 自花授粉后果实坐果率统计 梨是蔷薇科中自交不亲和物种[16],自花花粉可以在柱头上萌发,但是很少能生长到花柱底部[17-18]。本研究通过观察不同品种自花花粉管在花柱内停止生长的位置差异发现,许多品种花粉萌发后生长很短或者刚能穿过柱头就完全停止生长,表现为强自交不亲和性,而大多数品种自花花粉管生长停止在花柱长度的2/3之前,这与前人的报道一致[17]。然而,仍有少量花粉管能够生长至花柱基部,表现为弱自交不亲和性。每个花粉管数量在花柱中的分布从柱头到基部越来越少,且绝大多数花粉管最终在花柱内停止生长,花粉管末端形态结构改变。有活体试验表明,不亲和性花粉管生长受抑制前,有弯曲、先端膨大变形等现象,而异花授粉的花粉管没有此现象[10]。也有研究表明,自花授粉的花粉管在花柱内都有“快—慢—快”的动态生长变化过程[10]。在不同的梨品种中花粉管生长受到抑制时,花柱的发生部位及变化不同,徐义流等认为可能是自交不亲和基因时空表达的差异造成的[19]。也有研究表明,梨花柱中S糖蛋白的含量具有品种特异性和组织差异性,即品种不同,花柱中S糖蛋白的含量不同,并且S糖蛋白在花柱中分布不均匀导致花粉管停止生长[12,20]。梨的自交不亲和性主要表现在自交授粉花柱中花粉管的生长受到抑制,这与S糖蛋白有直接的关系,S糖蛋白的核糖核酸酶活性可通过降解花粉管RNA来抑制花粉管生长[21]。活体和离体试验均表明,自交不亲和性强度与花粉管长度呈负相关[11-12]。从而分析认为,由花粉管生长的长度可以判断自交不亲和强度,自交花柱中花粉管生长长度占花柱的比例将自交不亲和强度分为强、中和弱3类。在本研究中256个品种中,自交不亲和性强度参差不齐,强、中和弱频度分布也有很大差距。在离柱头近的地方S-RNase浓度可能较高,因此自交不亲和性为强的频度分布占多数,为梨不同品种花柱内S糖蛋白总量及其花柱内的分布提供一定的理论基础。S糖蛋白在花柱内是否具有梯度分布和怎样的分布有待进一步验证。本研究同时为后期深入研究自交不亲和性强度具体机制提供了充足的基础数据。 笔者通过对大量自花授粉花粉管生长状态研究,来初步判断筛选自交亲和性品种。发现金坠、秋荣和晚秀花粉管长到了花柱最底端,自交不亲和强度R值为100.00%。进一步自花套袋调查了坐果率,发现金坠和秋荣确实为自交亲和性品种。然而遗憾的是,晚秀在鉴定花粉管过程中极可能是自交亲和性品种,但最终自花套袋未结实。同时还发现,自交不亲和强度呈现中和强的品种,在自花套袋试验中也不结实。因此,这样的结果验证了笔者通过自花花粉管生长状态来初步筛选自交亲和性品种的必要性和可靠性,大大减少了盲目调查坐果率的工作量,并且丰富了自交不亲和强度R值数据,为后期进一步研究自交不亲和机制提供了充足的数据及一定的理论基础。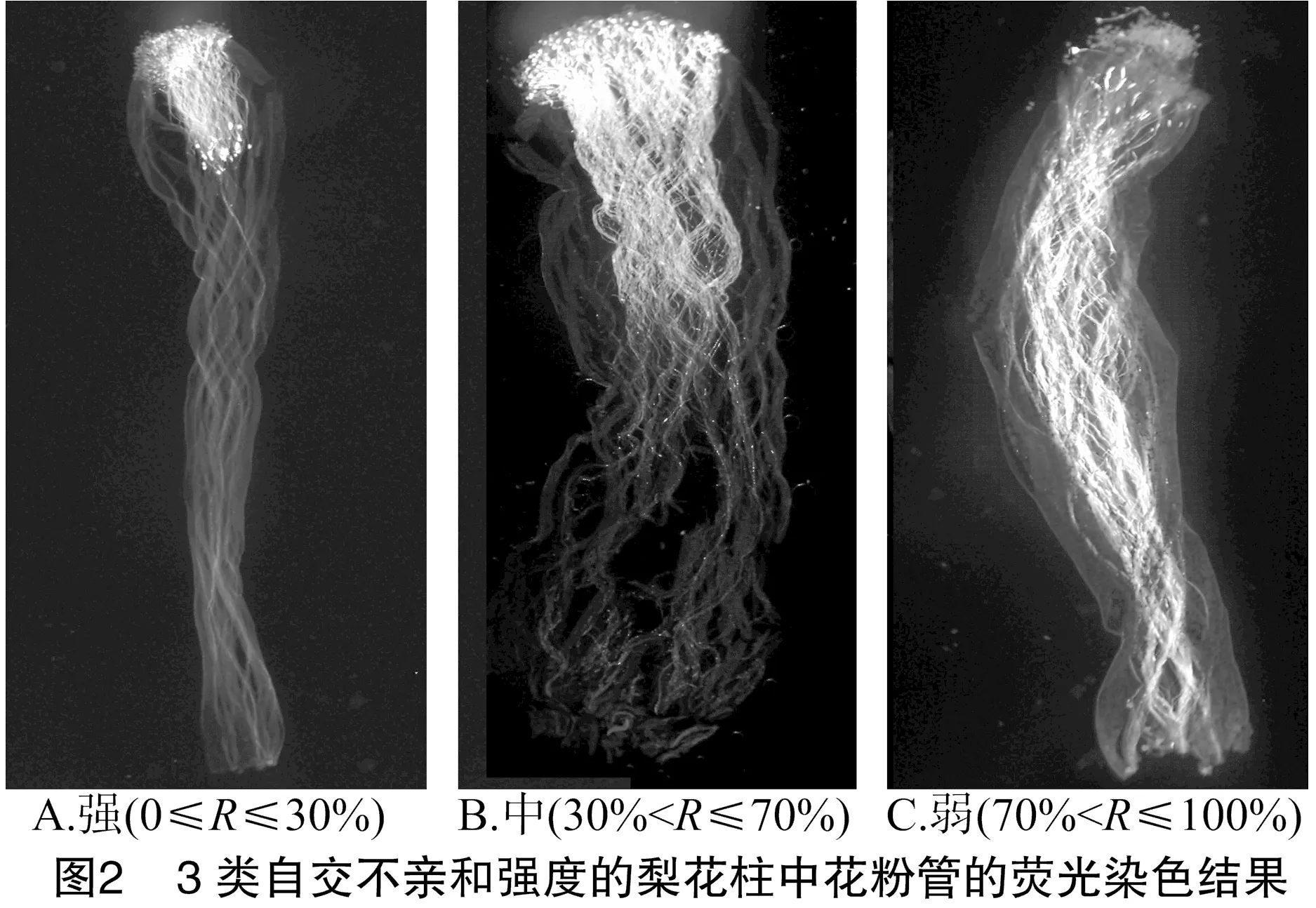
2.3 自交不亲和强度R值分析
2.4 不同梨品种自交不亲和性强度R值频度分布


2.5 自花授粉坐果率统计调查
3 讨论
