湿地松高产脂家系产脂量与胸径结构差异及其相关关系
2018-07-14许业洲杜超群范邦海方丽丹涂俊杰雷清虎李世升
许业洲,杜超群,范邦海,袁 慧,方丽丹,刘 星,涂俊杰,雷清虎,李世升
(1.湖北省林业科学研究院,湖北 武汉 430075;2.荆门市彭场林场,湖北 荆门 448269;3.黄冈师范学院,湖北 黄冈 438000)
湿地松Pinus elliottii原产美国东南部,我国于20世纪30年代开始引种栽培,因其具有生长快、适应性强、产脂量高等优势,现已发展成为我国南方低山丘陵地区的主要造林树种和重要的工业原料林树种。松香、松节油是我国出口创汇的主要林副产品之一,在松树人工林经济收益中占重要比重,高产脂优良资源的引种与选育自然成为其经营与科研的基础性工作内容。早在20世纪40年代美国林务局就开始湿地松产脂力的遗传改良及选育工作,对8年生人工授粉的子代产脂量的测定试验结果表明,湿地松高产脂力的性状具有显著的遗传性(狭义遗传力估计为45%~49%,广义遗传力估计为67%~90%)[1]。另外,美国还对湿地松和欧州赤松Pinus sylvestris天然林、印度对喜马拉雅长叶松Pinus roxburghii、西班牙和葡萄牙对海岸松Pinus pinaster、奥地利对欧洲黑松Pinus nigra等开展了产脂力的研究与高产脂类型的选育,均表明可通过遗传改良和选择提高松脂产量[2]。
我国近年来在湿地松[3]、马尾松[4-5]Pinus massoniana和思茅松[6]Pinus kesiya等主要采脂树种上开展了产脂力的遗传测定和相关因子分析工作,张建忠等[7]对15~16年生的17个湿地松高产脂半同胞家系的产脂量和生长量的遗传力进行了估算,指出湿地松高产脂家系的产脂量和生长量均处于较高遗传水平,且两者之间均达到极显著相关;庄伟瑛等[8]开展了16年生自选湿地松高产脂家系的产脂量研究,分析了家系间、家系内单株间产脂量的变化动态,得出湿地松家系产脂量遗传力高达0.9,并认为林木生长、形态特征对产脂量有显著影响。另外,庄家尧[9]、王光仁[10]、朱国发[11]等研究分析了湿地松胸径(径阶)、树高、冠幅等林木生长因子对产脂量的影响及其相关关系,均显示出湿地松产脂量与林木生长量有着紧密的相关性,生长势较优的林木产脂量较高。
本研究利用18年生湿地松高产脂家系保存林的单株测定数据,开展了家系产脂量遗传变异分析及其参数估算,并探讨了不同家系的胸径结构差异及其对产脂量的影响,建立了单木产脂量与胸径的线性关系,试图从测定和选择群体相结合的层面来分析不同家系产脂量的差异及其变化特性,为优良家系的选择及生产性人工林产脂量的评估提供参考。
1 材料和方法
1.1 试验地概况
试验地位于湖北省荆门市彭场林场,地处鄂中 腹 地 丘 陵 地 带,112°4′~ 112°14′E,30°35′~30°42′N,以丘陵岗地为主,海拔47 ~100 m,属北亚热带季风气候区,年平均气温15.9~16.2 ℃,年均积温5 050~5 100 ℃,无霜期约260 d,年降水量1 100~1 300 mm,全年气候温和,雨量充沛。林地土壤主要为第四纪母质发育的黄棕壤,土层深度在1 m以上,质地较粘,肥力中等,pH值5.6~6.8。
1.2 材料来源
试验材料为从美国引进的14个湿地松高产脂家系(种子由中国林业科学研究院提供),以彭场林场自选的高产脂混合家系为对照(见表1,对照编号为15)。1994年春用1年生实生苗造林,造林地为平缓坡地,全垦整地,各家系随机集中成片栽植,初植密度1 665 株/hm2,各家系造林面积0.02~ 0.10 hm2不等,保存林总面积0.80 hm2。幼林林粮间作,按常规抚育管理,10年生时进行强度约20%的下层疏伐抚育,现存林分密度约1 300 株/hm2。

表 1 各家系生长因子及产脂量数据简单统计Table 1 Data of growth factors and resin production of different families ofPinus elliottii
1.3 试验数据与分析
2014年对试验林进行每木调查,按常规测树方法实测胸径、树高、冠幅、枝下高,按采脂技术规程下降法“V”字型分别于7月、8月、9月对每株活立木进行产脂量测定,割脂高度为1 m左右,侧沟夹角70~90 °,深度达木质部0.3 cm,割面负荷率45%~50%,每3 d加割1刀,分单株称产脂重量,以3个月产脂量之和为单株产脂量指标进行统计分析。共计取得15个家系(包括对照)770株林木的生长因子和产脂量数据(见表1)。单株材积按照湖北省湿地松二元材积计算公式计算:
V=6.714 156×10-5×H0.7908×D1.9955。
用Excel、SAS、ForStat等软件进行试验数据的统计分析。
2 结果与分析
2.1 产脂量变异分析及遗传参数估算
试验材料平均单株产脂量为2.06 kg,变异范围为0.19~5.22 kg,变异系数为41.93%。按0.5 kg将单株产脂量分为10个等级,各产脂量等级的株数比例呈正态分布(偏度0.554 9,X2值5.613 9,小于X2的概率0.585 5)(图1),单株产脂量在1.5~2.0 kg的株数最多,占总株数的45.8%,超过84.2%的单株产脂量在1.0~3.0 kg之间,大于3倍标准差的株数比例为21.9%,有124株单株产脂量超过3.0 kg。

图1 产脂量数据分布情况Fig.1 Data distribution of resin production
表1中各家系平均单株产脂量的变动幅度为1.57~2.27 kg,与单株产脂集中分布的范围类似。各家系产脂量平均变异系数为44.2%(36.8%~48.6%),而家系间平均值的变异系数为14.5%。经方差分析,各家系间单株产脂量存在极显著差异(Pr>P<0.01)(见表2)。引进高产脂家系平均单株产脂量为2.11 kg,仅比对照大2.4%,其中只有7个家系单株产脂量超过对照,平均比对照大27.3%。

表 2 产脂量方差分析Table 2 AVOVA for resin production
分别以单株产脂量实测值和用线性函数估算的理论值进行家系产脂量遗传参数的估算(见表3),结果表明,家系产脂量遗传力均超过0.8,其理论值更接近0.9,而单株遗传力均占家系遗传力的一半以上,这与美国林务局[1]及庄伟瑛[12]等人的研究结果较一致,进一步说明产脂量受较高遗传效应控制,可进行有效的选择,其遗传增益达20%以上。

表 3 产脂量遗传参数估算Table 3 Estimates of genetic parameters for resin production
2.2 不同家系径级结构及其产脂量的差异
不同家系平均胸径变化幅度为17.8~20.1 cm,差异达到极显著水平(Pr>F=0.005 5)。将所有单株胸径(12.8~28.0 cm)按2 cm分级,各径级株数比例呈正态分布(偏度0.268 3,>X2的概率为0.699 0),但各家系的径级数量及其分布、相同径级家系数量及其株数与分布比例均存在明显差异(见表4),仅有2个家系在所有径级中均有分布,大多数家系(≥13个)分布在14.0~26.0 cm的6个径级中,而家系4的分布径级数量最少,仅为16.0~24.0 cm的4个径级,除在这4个径级所有家系均有分布外,分布在径级26~28 cm的家系最少(3个);按株数统计,整体林木中80.99%分布于14.0~22.0 cm的4个径级中,在最大径级(26.0~28.0 cm)的分布株数最少(5株),其次为较大径级(24.0~26.0 cm)的25株和最小径级(12.0~14.0 cm)的32株。

表 4 不同家系不同径级的林木数量Table 4 The number for different diameter-classes of different families
各径级平均胸径和平均产脂量的变动范围分别为13.5~26.8 cm和0.91~3.71 kg,在各径级内的平均变异系数分别为2.79%和31.42%。从图2中可以看出,胸径生长量在14.0~16.0 cm径级中变异系数最大,为3.80%,随后呈现出随着径级加大而逐渐减小的趋势;单株产脂量在最小径级(12.0~14.0 cm)的变异系数最大,随着径级增大逐渐减小,20.0~22.0 cm径级后基本趋于稳定。从方差分析结果看(见表5),相同径级不同家系间胸径差异除18.0~20.0 cm径级达到显著水平外,其它均无显著差异,这可能与不同家系在该径级的数量分布差异较大(4.80%~61.50%)有关;家系间产脂量差异除最小径级(12.0~14.0 cm)和较大径级(24.0~26.0 cm、26.0~28.0 cm)不显著外,其它径级均呈现极显著差异。这一结果表明,即使在相同径级或者胸径差异不显著时,不同家系间的产脂量依然表现出显著差异,进一步说明产脂量受到较强的遗传控制,但当林木在群体中处于竞争两极时(被压木或优势木),这种遗传控制可能受到一定程度的抑制或者说更多地受到林木个体生长状态的影响而致使其差异缩小到不足以显现。
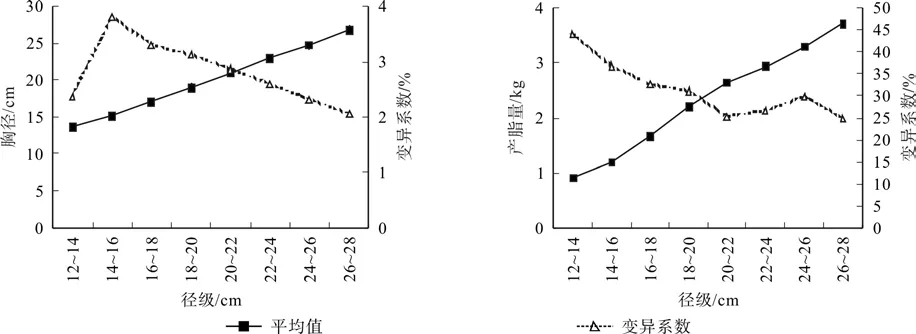
图2 胸径和产脂量随径级的变化趋势Fig.2 Variation tendency of DBH and resin production with the change of diameter-classes
2.3 单株产脂量与生长因子间的相关关系
将单株产脂量与其生长因子进行协方差分析(一元线性回归),模型达到极显著水平(R2=0.552 8,Pr>F<0.01)。从表6中可以看出,生长因子F值从大到小的排序为胸径>材积>树高>冠幅>枝下高,而胸径的F值是其它生长因子的8倍以上,说明胸径对单株产脂量的影响远大于其它生长因子。除枝下高协方差不显著外,树高和冠幅协方差均达到显著水平,而胸径和材积达到极显著水平。

表 5 不同径级家系间胸径及产脂量方差分析Table 5 AVOVA for DBH and resin production in different families of different diameter-classes
为了进一步弄清各生长因子对产脂量的影响,将产脂量与除材积外的生长因子进行逐步回归分析,结果表明(表7),回归模型达极显著水平(R2=0.547 7,Pr>F<0.01),胸径回归系数和偏相关系数均为最大值,而且分别比其它生长因子大3倍和9倍以上。一般认为,胸径较大的林木具有明显的生长优势和健壮旺盛的生命特性,胸径是林木生长因子中最根本和最关键的因子,因此也对产脂量有着极大影响,两者之间存在显著相关关系。

表 6 产脂量与生长因子的协方差分析Table 6 Covariance analysis of resin production and growth factors
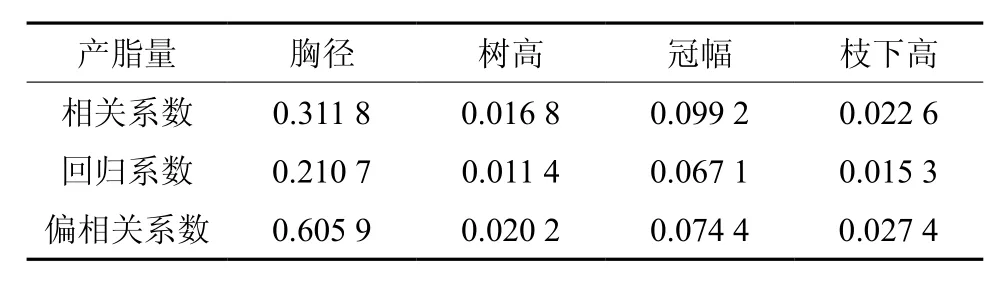
表 7 产脂量与生长因子回归分析Table 7 Regression analysis of resin production and growth factors
2.4 产脂量与胸径的线性关系
以单株产脂量(因变量y)与其胸径(自变量x)进行一元线性回归,其函数关系式为y=0.224 3x-2.112 4,R2=0.544 2,P<0.01,其相关系数和回归系数检验均达到极显著水平(见表8)。对该函数进行适应性检验,结果表明理论值与实测值差异不显著,可作为生产中单株产脂量的估测模型。
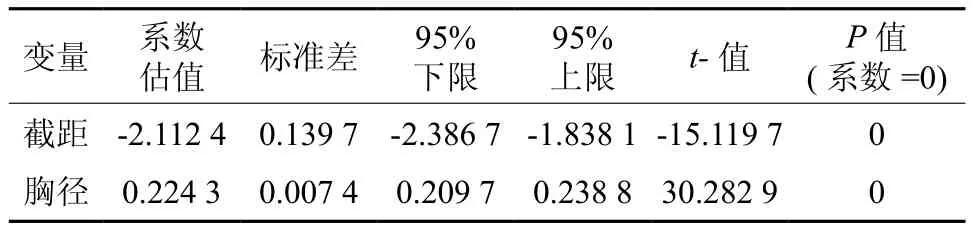
表 8 产脂量与胸径回归系数Table 8 Regression coefficient of resin production and DBH
进一步分别各家系进行单株产脂量与胸径的一元线性回归(表9),除4号家系外,其它家系的回归相关系数为0.512 1~0.828 0,且均达到极显著水平(P<0.01)。可能由于4号家系保存株数较少,其回归相关系数仅0.343 2,但其线性函数同样达到显著水平(P=0.027)。与总体线性函数相比,各家系单株产脂量与胸径的函数参数相近,其平均系数和截距仅分别相差3.3%和6.2%。

表 9 不同家系单株产脂量与胸径的线性函数Table 9 Linear function of resin production and DBH in different families
从图3可以看出,各家系直线斜率存在一定差异,变化幅度为0.162 5~0.320 2,平均斜率为0.231 8,变异系数18.8%,各家系单株产脂量与胸径线性函数的总体趋势基本一致,不同家系单株产脂量随着胸径增大而逐渐增大,但家系间并非理想的平行线关系,而是呈现出部分交叉重叠的放射状直线簇,表现出不同家系随着胸径增长的增幅存在一定差异和波动,并非在较小径级产脂量较高的家系就一定在较大径级中同样具有较高产脂量,这与上节不同径级不同家系产脂量差异的研究结论一致。
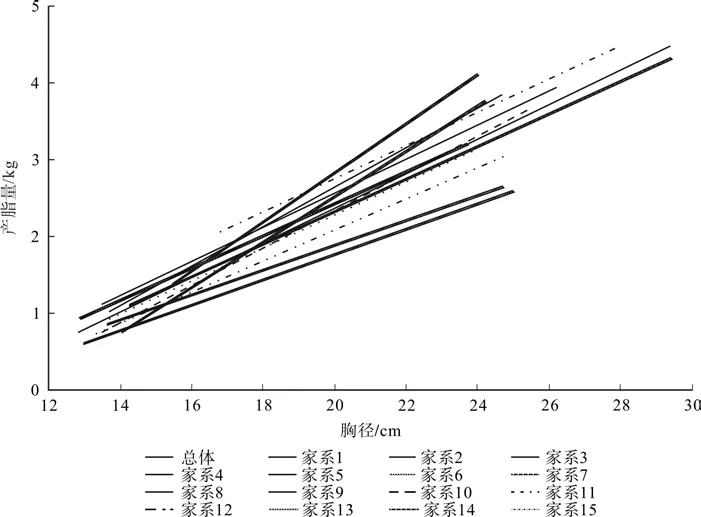
图3 各家系单株产脂量与胸径的线性关系Fig.3 Linear relation of resin production and DBH in different families
3 结论与讨论
本研究对18 a生15个湿地松家系(包括对照)产脂量的变异情况进行了统计分析,结果表明家系间产脂量存在极显著差异,家系遗传力为0.809 6,而单株遗传力占家系遗传力的50%以上,均达到较高水平。这一结论略高于张建忠[7]对同一批材料江苏试验点15~16 年生的家系遗传力(分别为0.674 8和0.693 0),但略低于庄伟瑛[8]16年生自选湿地松高产脂家系子代测定林家系遗传力0.90,而与美国研究的湿地松产脂遗传力0.80的结论较为一致[1,12],本研究结论同样表明,湿地松产脂量受到较强遗传控制,通过家系进行高产脂育种材料的选择是十分有效的。但在本研究引进的14个高产脂家系中,仅有7个家系的产脂量高于本地对照,产脂量最高的家系仅比对照大31.6%,这与胡兴宜[13]和张建忠[7]得出的引进高产脂家系产脂量普遍高于本地对照的研究结论并不一致,这很可能与本研究选用的对照材料是以引进的美国优良家系经产脂量测定选择出的高产脂优良单株的混合子代有关。
尽管整体林分的径级结构呈现正态分布,但不同家系的径级分布,包括不同家系的径级数量和不同径级的株数均表现出较明显差异,这可能与不同家系的林木保存数量有关,但同时也体现出不同家系的生长特性和竞争状态具有一定的差异性。除最小径级(12~14 cm)和较大径级(24~28 cm)外,其它相同径级不同家系的产脂量均为极显著差异,这一结果表明不同家系产脂量可能更多受到处于竞争状态的中间群体的产脂量大小的影响。舒文波等[14]对马尾松人工林进行了不同年龄、不同径级产脂量变化特征研究,结果表明同一年龄不同径级的产脂量达到显著差异,且随着径级增大,产脂量相应增加,这与本研究产脂量随径级增大逐步增加的结论一致。
很多研究表明立木单株产脂量与其胸径、树高、冠幅、枝下高、材积等生长因子具有密切相关性,本研究结果也符合这一基本结论并进一步验证了湿地松单株产脂量与生长量之间的相关关系,其中胸径对产脂量的影响最大,两者协方差F值超过其它因子8倍以上,逐步回归偏相关系数也超过其它因子9倍。王光仁等[10]研究表明40年生湿地松产脂量与胸径和树高均存在极显著相关,张建忠等[7]研究认为高产脂湿地松的生长量(胸径、树高、材积)与产脂性状之间都达到极显著相关,而本研究显示出胸径对单株产脂量具有更为突出的影响,这与赖标祯[15]对马尾松产脂量与树高、胸径、材积的关联度分析,舒文波等[14]对20 年生马尾松林产脂量与胸径、树高、枝下高和树冠比的相关分析,黄帆等[4]对32 年生马尾松产脂量与胸径、树高、冠幅的相关性分析以及翁海龙等[6]对266株思茅松高产脂优树的主要性状做相关性分析等研究结论较为一致,均表明胸径是生长性状中与产脂量相关最为密切的因子。因此,在高产脂优良家系及其生长指标的多性状联合选择过程中,可实现胸径与产脂量的联合选择,胸径生长量越大,产脂量也就越高。
本研究建立单株产脂量与胸径的一元线性回归函数,其相关系数和回归系数检验均达到极显著水平,与朱永安等[11]建立的24年生湿地松产脂量与直径一元线性方程相比,两者的系数和截距十分相近(见表10)。如果分别利用以上2个一元线性方程对本试验材料进行单株产脂量的估算,两者估算值的统计参数也很接近,后者总体估算平均值偏高,这可能与其树龄较大而产脂量更高有关,相差比例为9.8%。在实际产脂量的测定中,由于采脂方式和操作手法的差异,以及松脂承接与收集中的诸多问题,造成测量数据和试验结果的较大误差,而胸径是林木生长因子中最容易也更精确的测量指标,建立两者的函数关系进行产脂量的科学预估,对湿地松高产脂人工林的经营管理具有重要的指导意义。

表 10 与24年生湿地松产脂量和胸径线性函数的比较Table 10 Comparison of linear function of resin production and DBH withPinus elliottiiin 24-years-old
