Comparative study of responses in the brown algaeSargassum thunbergiito zinc and cadmium stress*
2018-07-11Fang吕芳DINDGang丁刚LIUWei刘玮ZHANDongmei詹冬梅WUHaiyi吴海一GUOWen郭文
LÜ Fang (吕芳) DIND Gang (丁刚) LIU Wei (刘玮) ZHAN Dongmei (詹冬梅) WU Haiyi (吴海一) GUO Wen (郭文)
1Marine Biology Institute of Shandong Province,Qingdao 266104,China
2Qingdao Macroalgae Engineering Technology Research Center,Qingdao 266104,China
AbstractHeavy metal pollution in aquatic system is becoming a serious problem worldwide. In this study, responses ofSargassum thunbergiitodifferentconcentrations (0, 0.1, 0.5, 1.0 and 5.0 mg/L) of zinc(Zn) and cadmium (Cd) exposure separately were studied for 15 days in laboratory-controlled conditions.The results show that the Specific growth rates increased slightly under the lower Zn concentration treatment(0.1 mg/L) at the fi rst 5 d and then decreased gradually, which were significantly reduced with the exposure time in higher Zn concentrations and all Cd treatments compared to respective control, especially for 1.0 and 5.0 mg/L Cd. Chlorophyllacontents showed significant increase in 0.1 mg/L Zn treatment, whereas the gradually reduction were observed in the other three Zn treatments and all Cd treatments. The oxygen evolution rate and respiration rate presented distinct behavior in the Zn-treated samples, but both declined steadily with the exposure time in Cd treatments. The P/R value analyses showed similar variation patterns as chlorophyll a contents. Real-time PCR showed that lower Zn concentration (0.1 mg/L) increased mRNA expression ofrbcL gene, whereas higher Zn concentrations and Cd reduced therbcL expression. Taken together, these fi ndings strongly indicate that Zn and Cd had different effects onS.thunbergiiboth at the physiological and gene transcription levels, the transcript level of photosynthesis-related generbcL can be used as an useful molecular marker of algal growth and environment impacts.
Keyword: Sargassum thunbergii; heavy metal stress; chlorophyll; photosynthesis rate; respiration rate;rbcL gene
1 INTRODUCTION
Heavy metal pollution in aquatic system has become a global environmental pollution problem.This fi eld has captured more attention globally and this is due to the fact that, these pollutants cannot be decomposed through biological degradation and have the ability to accumulate in the environment, and eventually become persistent pollutants (Asaolu and Olaofe, 2005; Olowu et al., 2010). Macroalgae are important primary producers in marine ecosystem,which not only can remove nutrients N, P and other organic pollutants, but also can absorb heavy metals in the ocean, so it can be applied to the remediation of heavy metal pollution in marine ecosystem (Volesky and Holan, 1995).
To improve the understanding of heavy metals absorption by macroalgae, a number of attempts have been made to monitor physiological changes induced by heavy metals contamination on the macroalgae(Amado Filho et al., 1997; Xia et al., 2004; Saleh,2015; Costa et al., 2016a, b, 2017). These results have shown that heavy metals affect macroalgal growth rates and impairs important cellular processes like photosynthetic electron transport, photosynthesis,and respiration (Gouveia et al., 2013; Costa et al.,2016a, b). And signifi cative differences in the physiological and metabolic processes responses under different metals stress were also observed(Volesky and Holan, 1995; Santos et al., 2015). It is well established that, Zn is an essential micronutrient for catalyzing enzymatic reactions in plants but when present in excess can become extremely toxic and threshold survival (Santos et al., 2015). Comparatively,Cd is not an essential nutrient and it is one of the heavy metals that are known to generate toxicity even at a very low concentration (Hall, 2002), which indicating the activation of different mechanisms that promote the uptake and toxicity of Zn and Cd.
Photosynthesis is a fundamental process common to all ecosystems, and environmental factors have been shown to significantly inf l uence this process.Several studies already demonstrated the photosynthetic responses to heavy metal stress of macroalgae (e.g. Mamboya et al., 1999; dos Santos et al., 2012), however, little information exist discussing the expression of photosynthetic genes to increased heavy metal concentrations. The latter is thus highly desirable to fully understand the photosynthetic responses of macroalgae associated with the heavy metal stress process.
It has been proved that someSargassumspecies had the high bioaccumulation capacity of heavy metals and can serve as biosorbent agent or bioindicator for heavy metal contaminated marine environment (Jothinayagi and Anbazhagan, 2009;Patrón-Prado et al., 2011; Costa et al., 2016a). This distinctive phenomenon of thoseSargassumspecies had provided excellent models for bioremediation of heavy metal pollution studies.Sargassumthunbergii(Mert.) O’ Kuntze is an economically important seaweed along the coast regions of China, which not only can be used for the extraction of alginate,mannitol, iodine and fucoidan in industry, but also an ideal biological material for marine ecological environment reparation. Our previous studies have proposed thatS.thunbergiihas a high ability to accumulate the Zn and Cd (Wu et al., 2010). In this study we made further investigations for the response ofS.thunbergiito different Zn and Cd concentrations exposure. Their growth, pigment content,photosynthetic oxygen evolution and the relative expression of large-subunit ribulose-1, 5-bisphosphate caboxylase / oxygenase (rbcL) mRNA were investigated in indoor seawater culture systems. It is hypothesized that Zn and Cd had different effects onS.thunbergiiboth at the physiological (algal growth,chlorophyll and photosynthesis) and gene transcription(photosynthesis-related genes:rbcL) levels.
2 MATERIAL AND METHOD
2.1 Algal materials and sample collection
S.thunbergiiwere sampled from intertidal zone of Taiping bay (36°05′N, 120°35′E), Qingdao, China in April of 2014. The algal samples were rinsed with plenty of autoclaved seawater and brushed by a soft brush to remove the surface microbial and epiphytic organisms. Then the algal thalli were maintained in a seawater aquarium at 15°C, illuminated by a light of 1 000 lx (light was measured with a Quantitherm Light Meter, Hansatech, England) and aerated with normal air for 5 days, and then individuals with robust growth, consistent confi guration and length in 4-5 cm of the individuals were selected for experiments.
The heavy metal solutions were prepared by using analytical grade CdCl2·2.5H2O and ZnSO4·7H2O salt respectively. Concentration of metal ions were 0.1,0.5, 1.0, 5.0 mg/L, and the natural seawater with no metal addition as control groups. Proposed concentrations were based on our previous analyzed data of Wu et al. (2010). The experiments were performed in 3 000 mL beakers, each containing 2 000 mL solution and each treatment group comprised of 3 replicates of 15 strains of algae (~10.0 g FW).Culture medium were replaced once a day. The treatments were put into a Plant Incubator HP1000G(Wuhan Ruihua Instrument & Equipment Co., Ltd.,China) at a temperature of 15°C, continuous aeration,and 12 h light/12 h dark cycle (light on 08:00 to 20:00,the intensity was 3 000 lx (f l uorescent lamps, FSL T8 30 W/765)).
The following measurements were performed using the three parallel experiment samples which were randomly sampled at 0, 3, 5, 10 and 15 days after cultivation from each groups.
2.2 Specific growth rate measurements
The formula of Specific growth rate (RSG) was
5. Taking my husband for granted. Just as I find it easily to overlook(,) the chores done by my husband, it’s easy for me to forget to appreciate his many virtues13 and instead focus on his flaws. For example, although I find it hard to resist using an irritable tone, my husband almost never speaks harshly, and that s really a wonderful trait. I’m trying to stay alert to all the things I love about him, and let go of my petty annoyances14. This is easier said than done. I’ve found that working to keep my resolution to Kiss more, hug more, touch more is an effective way to help me stay in loving, appreciative frame of mind.

whereWtwas the fresh weight (g) of algae at the middle or end in experiment,W0was the fresh weight(g) of algae at the beginning of the experiment,twas the culture time in days (Lignell and Pedersén,1989).

Table 1 Real-time quantitative PCR primers
2.3 Pigment extraction and light absorption analysis
Chlorophyll-a(chla) concentration was measured using the spectrophotometric assay based on the method of Jeffrey and Humphrey (1975). The algal samples were lyophilized and ground to a fi ne powder in liquid nitrogen, and then 8 mL of 80% acetone solution was added and then kept in darkness at 4°C for 12 h. The acquired mixture was centrifuged at 10 000×gand 4°C for 10 min, the supernatant was adjusted to 10 mL for further analyses. The extractions were then analyzed with a spectrophotometer with 80% acetone solution as the blank. Absorbance was recorded at 630,664 nm. The contents of chlawas calculated following formula Ca=11.47OD664- 0.40OD630, and was standardized to fresh mass.
2.4 Oxygen evolution measurements
The photosynthetic oxygen production was determined using a Clark-type electrode (Chlorolab-3,Hansatech, England). All measurements were made at ambient temperature of 20°C controlled by a thermostatic water bath. Oxygen production rates were measured under predetermined saturating irradiances (300 μmol/(m2·s)). The rates of oxygen evolution and respiration were standardized to fresh weight biomass (nmol O2/(min·g) FW), the value of P/R was obtained by the ratio of photosynthetic rate to respiration rate.
2.5 Semi-quantitative RT-PCR analysis of rbc L gene expression
Total RNA was extracted using the E.Z.N.A.TMPlant RNA Kit (Omega Biotek). Quality and quantity of extracted RNA from all samples was confi rmed by both agarose gel visualization and spectrometry. And then MMLV reverse transcriptase (Promega) was used to synthesize single-strand cDNA with the DNase I (Promega)-treated total RNA as template and oligo (dT) primer. The reaction was incubated at 42°C for 1 h, terminated by heating at 95°C for 5 min, and subsequently stored at -80°C.
Following the parameters of real-time PCR oligonucleotide primer design (Ferreira et al., 2006),primers were designed for 18S rRNA and therbcL gene (Table 1), with Primer 5 based on sequences in the GenBank database.
Real-time PCR assays were performed on CFX96 real-time PCR detection system (Bio Rad, USA) in a 25 μL reaction volume containing 12.5 μL of SYBR®Premix Ex Taq (Tli RNaseH Plus) (2×) (TaKaRa,Japan), 2 μL template (<100 ng) and primers at a concentration of 200 nmol/L. The PCR programs was carried out at 95°C for 10 min, followed by 40 cycles of 95°C for 10 s, 58°C for 30 s and 72°C for 30 s. At the end of each reaction, melting curve analysis of PCR products was taken in order to insure the Specificity of the amplifi cations. The expression level of therbcL gene was calculated using the formula: 2-∆∆Ct.
2.6 Statistics
All values were calculated as an average of three independent experiments (n) and are presented as mean±standard deviation. Statistical analysis was performed with Student’st-test using software SPSS 13.0 for window, andP<0.05 was set as the level of statistical signifi cance.
3 RESULT
3.1 Specific growth rate
The Specific growth rate ofS.thunbergiidid not change significantly in the control group during the 15 d exposure to Zn, while the growth rate raised at the fi rst 5 d in the low concentration group (0.1 mg/L),and then decreased gradually, Whereas, when applied Zn concentration increased to 0.5 mg/L, the growth rate has been decreasing during the whole exposure period, and more noticeable at the higher applied Zn concentration (1.0 and 5.0 mg/L), reaching values significantly lower than the control group (P<0.05)until the end of the 15 d culture.
WhenS.thunbergiiexposed in different concentrations of Cd solutions, the growth rate of each experimental group decreased gradually with the exposure time which compared to control. As presented in Fig.1, growth rate decreased as Cd applied concentrations increased, especially the negative growth in higher concentration groups (1.0 and 5.0 mg/L).

Fig.1 Effects of different concentrations of Zn and Cd (0.1,0.5, 1.0 and 5.0 mg/L) on Specific growth rate of S.thunbergii for 15 days
3.2 Light absorption analysis
WhenS.thunbergiiexposed in Zn solution, there was a significant increase in chlacontent under 0.1 mg/L Zn than their respective control, whereas the gradually reduction were observed in the other three experimental groups with 0.5, 1.0 and 5.0 mg/L treatments.
However whenS.thunbergiiexposed in Cd solution, the chlacontent of each experimental groups were decreased gradually with the time of exposure. As presented in Fig.2, there were significant differences between the different concentration groups (P<0.05), chlacontent decreased as Cd applied concentrations increased.
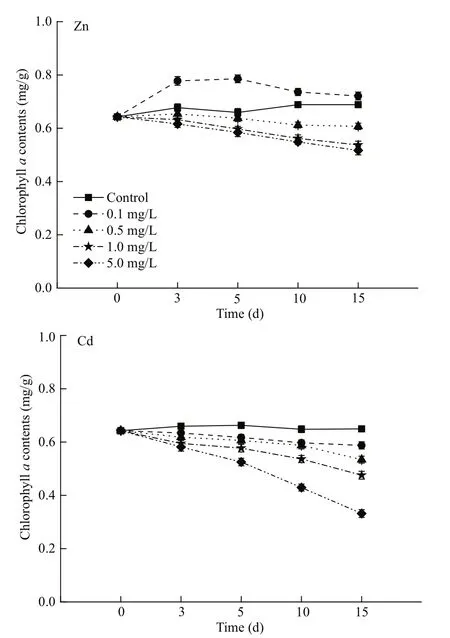
Fig.2 Effects of different concentrations of Zn and Cd (0.1,0.5, 1.0 and 5.0 mg/L) on chl a content of S. thunbergii for 15 days
3.3 Rate and effciency of photosynthetic oxygen evolution
Compared to control group, heavy metals stress caused a significant change in the oxygen evolution rate and respiration rate, as shown by the oxygen exchange parameters (Fig.3). whenS.thunbergiiexposed in Zn solution, the oxygen evolution rate increased slightly in the low concentration group(0.1 mg/L), but declined steadily in the other higher concentration groups; the respiration rate showed distinct behavior in which no significant difference in lower concentration group (0.1 mg/L), Whereas,when applied Zn concentration were 0.5 and 1.0 mg/L,the values remained relatively constant in the fi rst 5 d and then tended to decrease at different rate.
While, whenS.thunbergiiexposed in Cd solution,both the oxygen evolution rate and respiration rate for all the groups declined steadily with increased time of exposure.
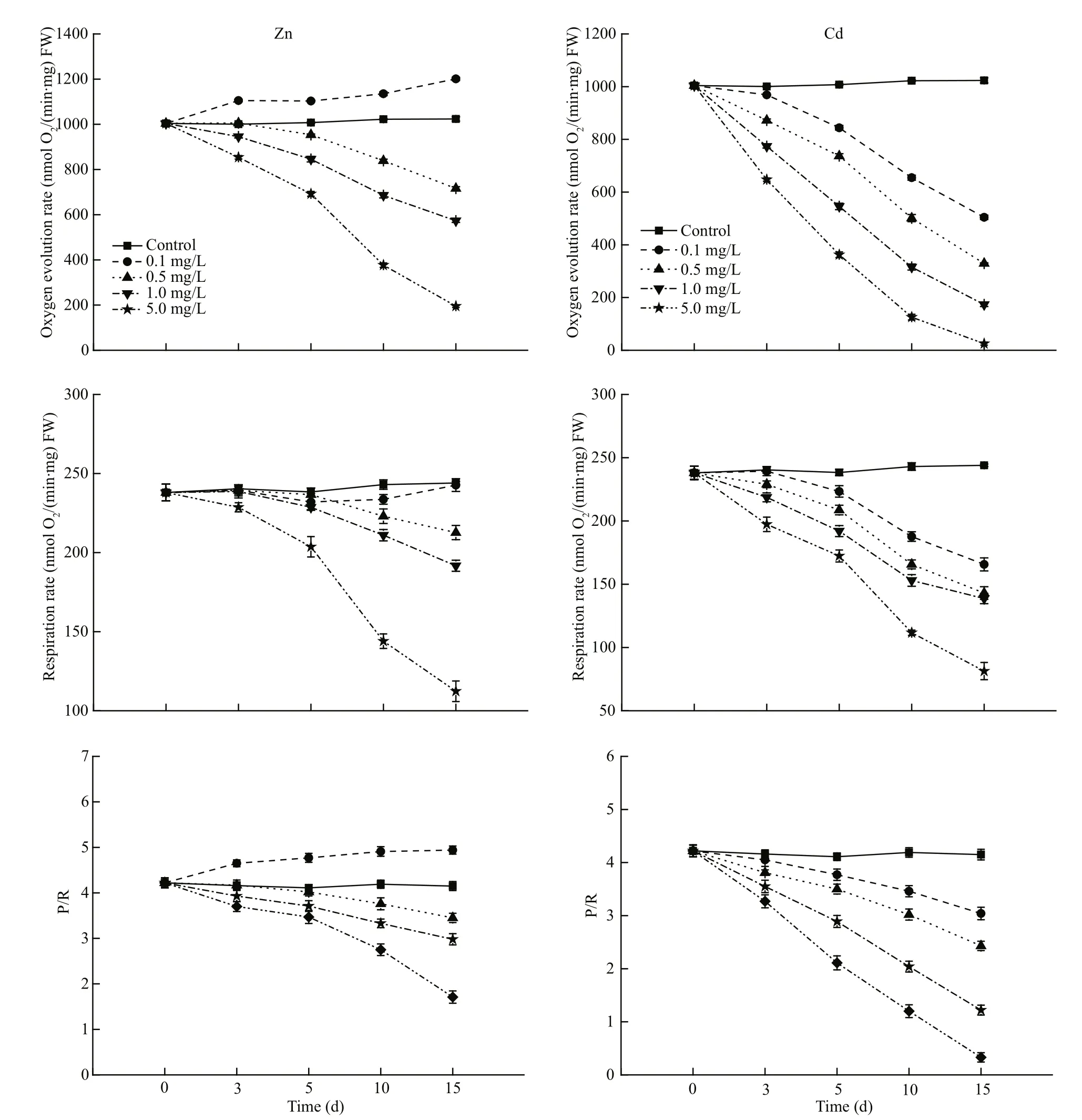
Fig.3 Effects of different concentrations of Zn and Cd (0.1, 0.5, 1.0 and 5.0 mg/L) on the oxygen exchange parameters(oxygen evolution rate, respiration rate and P/R value) of S. thunbergii for 15 days
We use the ratio of photosynthetic rate to respiration rate (P/R) to evaluate the metabolic activity during heavy metal exposure. This data did not change significantly in the control group during the experiment, which was slightly higher when exposure to lower concentration Zn (0.1 mg/L) than the control,Moreover, when applied Zn concentration increased to 0.5 mg/L, there was a significant decline in P/R values as exposure time and Zn ions applied concentrations were increased. However, P/R values significantly decreased of each Cd treatment groups,and Cd stress caused a progressive decline in the P/R values as Cd applied concentration increased.
3.4 rbc L gene expression

Fig.4 mRNA expression of rbc L at different time points during different concentrations of Zn and Cd (0.1, 0.5, 1.0 and 5.0 mg/L) treatments by RT-PCR analysisVertical bars represented the mean±s.d. (n=3).
The temporal expression of therbcL gene during heavy metal exposure was measured by semiquantitative RT-PCR (Fig.4). Our results showed that there was no significant difference inrbcL gene expression among the control group, but whenS.thunbergiiwere exposed in Zn solutions, the mRNA expression ofrbcL in lower concentration group(0.1 mg/L) was increased continuously with the exposure time, and it was up-regulated by 6.27-fold on day 15 compared with that at the beginning. While,in contrast, these values dropped gradually along with the exposure time in the higher concentration groups(0.5, 1 and 5 mg/L), and the higher Zn concentration,the greater decline values.
However, therbcL gene expression showed very different patterns when exposure in Cd solutions,rbcL transcript levels of all treatments dropped steadily along with exposure time, and reaching values significantly lower than the control group until the end of the 15 d exposure. Also correlation existed between metal concentration andrbcL gene expression, the higher Cd concentration treatment,the lower gene expression.
4 DISCUSSION
4.1 Effects of heavy metals on growth
Zn and Cd are the common heavy metal pollutants in aquatic environment, however, our experiments indicate that growth ofS.thunbergiishowed different patterns of variation during Zn and Cd treatments respectively (Fig.1). On one hand, Zn are required at low concentrations and can be toxic at higher concentrations, as shown forS.thunbergii, which was stimulated at free Zn concentrations 0.1 mg/L and inhibited at concentrations higher than 0.5 mg/L; on the other hand, Cd inhibited the growth ofS.thunbergiieven at low concentrations 0.1 mg/L and their inhibitory effect increased with increase in the concentration of metals. And the comparison of the results showed that a higher decline in the growth of algae was presented with Cd treatment than Zn treatment. Similar fi ndings were also documented in green algaeUlvalactuca, which their growth was more impaired with Cd treatment than the other tested metal ions (Pb, Cu , Zn) (Saleh, 2015). This observation could reflect the capacity of algae to tolerate the Zn ion compared to Cd. This different effect of these two ions maybe due to the fact that Zn was an essential minerals for catalyzing enzymatic reactions in algae, while Cd was nonessential element for macroalgal growth,development, and physiological processes (Xia et al.,2004) which can generate toxicity even when only applied at a very low concentration. The decrease in growth rates of algae may be related to the use of energy for activation of adaptation mechanisms and repair of damage induced by heavy metal stress.
It is worth noting that there was significant correlation with growth inhibition and exposure time,when more than 5 d especially 10 d treatment, the growth ofS.thunbergiiwas significantly inhibited,this indicates thatS.thunbergiihas a certain tolerance to Zn and Cd toxicity. However, even closely related species have been reported to exhibit different accumulation strategies for heavy metals. Amado Filho et al., (1997) reported the growth and Zn accumulation in six algae species under different levels of Zn, and stated that all six examined seaweeds died 3 days with 5 mg/L Zn. Among the tested algae,brown algaPadinagymnosporaandSargassum fi lipendulapresented the highest accumulation level of Zn. These results suggest that there is a need for careful choice of species for monitoring heavy metals programs.
4.2 Effects of heavy metals on photosynthesis
Toxic effects of heavy metal on plant reflected in the inhibition of photosynthesis, respiration and nitrogen fi xation, reduce the activities of enzymes,affect the metabolism and physiological process of algae (Xia et al., 2004; dos Santos et al., 2012).Chlorophyll is the main photosynthetic pigment, and its performance is closely related to the photosynthesis of plants, which reduction is an important indicator of plant senescence. Our data revealed marked changes in chlacontent ofS.thunbergiiafter exposure to Zn and Cd treatment (Fig.2). From data presented herein,it was noticed that slightly elevated in chlacontent under 0.1 mg/L Zn than their respective control, while 0.5, 1.0 and 5.0 mg/L Zn, caused reduction in chlacontent by 11.8%, 22.0%, and 24.9%. Whereas, under Cd treatments this decline was found to be 17.8%,26.7%, and 48.9% below the control forS.thunbergiirespectively. Our results, consistent with other heavy metal stress records in algae (Xia et al., 2004; Saleh,2015). The reduction in chlorophyll content could be associated with following reasons: (1) chlorophyll synthesis was inhibited. Alberte and Thornber (1977)found that the chlorophyll loss could be accounted for by reduction in the lamellar content of the lightharvesting chlorophylla/b-protein in stressed plants.Stobart et al. (1985) found that Cd inhibiting the formation of chlorophyll by reacting with essential thiol groups in both the protochlorophyllide reduetase protein and the enzyme(s) involved in the light dependent synthesis of 5-aminolaevulinie acid.However, Xia et al., (2004) thought that the decline in chlorophyll content after exposure to heavy metals could be related Mg and Fe defi ciency in the biosynthetic process of chlaor the inhibition of enzyme activity, i.e., photochlorophyllide reductase;(2) chlorophyll were degradable, this may be related to the increased activity of enzymes which promoted the decomposition of chlorophyll.
Zn and Cd treatments also had significant effects on photosynthesis ofS.thunbergiirelative to the control (Fig.3). Our data showed that oxygen evolution rates decreased by 50.7%, 67.8%, 83.1%,and 97.6% at 0.1, 0.5, 1.0 and 5.0 mg/L Cd exposure during 15 d. Whereas, this reduction was recorded to be 32.1%, 41.4%, 43.2%, and 66.7% for respiration rate under the above mentioned Cd concentrations. In contrast, the oxygen evolution rate increased by 17.3% after exposure to 0.1 mg/L Zn, the respiration rate remained relatively stable, and both were decreased at elevated Zn levels. Those data further confi rmed that high heavy metal concentration led to photosynthetic depression. In this study Zn and Cd treatments had a more apparent effects on oxygen evolution rate than respiration rate, which indicated that chloroplasts were more sensitive to Zn or Cd treatments, but further works would be needed to prove it.
P/R has been suggested as an effective parameter in evaluating the signifi cance of productivity measurements (Humphrey, 1975). The ratio is known to vary with many factors including growth phase(Ryther, 1955) and nutrient levels (McAllister et al.,1964), and this ratio is about 4:1 in the normal algae(Melis and Melnicki, 2006), In present study, the P/R ratio of theS.thunbergiiin the control group was about 4.1, showing a normal photosynthetic capacity.Under Zn and Cd treatment, P/R ratio followed similar variation patterns as the chlacontents, and during 15 d exposure, the P/R value is only about 1.74 and 0.31 respectively, which indicates that the photosynthesis ofS.thunbergiihas been greatly damaged.
4.3 Effects of heavy metals on rbc L gene expression
Rubisco is an important functional protein and energy storage protein in plants, and it is a key ratelimiting enzyme in the photosynthetic carbon assimilation process, which is important to photosynthesis research. It has been shown that the expression ofits coding generbcL for subunit was affected by the growth condition and environment(Pichard et al., 1993; Xu et al., 2012; Shao et al.,2014). In our study, mRNA levels ofrbcL decreased gradually with time after exposure to Cd, whereas increased steadily at 0.1 mg/L Zn treatment and decreased at higher concentrations. The changes inrbcL expression were consistent with those detected in photosynthesis rates, so we speculate that controlling the expression ofrbcL gene is one of the mechanisms of Zn and Cd effects onS.thunbergiiphotosynthesis, thereby affecting Rubisco synthesis,and that will eventually affect photosynthesis. Zn or Cd treatments on macroalgae could inf l uence photosynthesis partly at the genetic level.Nevertheless, photosynthesis is regulated at multiple levels, the inf l uence of heavy metals on photosynthesis cannot be deduced only according to the transcript levels, so it needs to be further studied in the future.
It has been shown that the expression ofrbcL gene in macroalgae can be inf l uenced by temperature,salinity, light, nutrients and other environmental factors. Xu et al. (2012) investigated therbcL gene expression ofUlvaproliferaunder various forms and intensities of stress (differing levels of salinity, light intensity and temperature), and found that the expression ofrbcL were higher under adverse circumstances except under conditions of desiccation.Shao et al. (2014) demonstrated that the mRNA levels ofrbcL in gametophytes ofSaccharinajaponicaexhibited diurnal variations: increased expression during the day but suppressed expression at night.This reminds us thatrbcL gene could be used as a molecular marker for diagnosis the growth of macroalgae, which evaluation response characteristics of macroalgae to environmental changes by the expression ofrbcL gene, this will provide a new explication for evaluation the environmental impacts on marine economic algae.
5 CONCLUSION
In sum, the results of the present study lead to the conclusion that heavy metals Zn and Cd had different effects on brown algaeS.thunbergiiand Cd has greater toxicity than Zn in terms of algal growth, chlaand photosynthesis. Analysis showed that low concentrations of Zn (0.1 mg/L) stimulated growth ofS.thunbergiiand increasing chlacontents, the oxygen evolution rate and therbcL gene expression, while higher Zn concentrations and all Cd treatments inhibited those compared to their respective control.Moreover, this growth inhibition was significant correlated with exposure time. Furthermore, our results provide additional evidence that both Zn and Cd may affect photosynthesis at the genetic level. The transcript level of photosynthesis-related generbcL variation was consistent with chlacontents and photosynthetic parameter P/R values, we supposed thatrbcL gene could be used as a molecular marker for diagnosis the growth of macroalgae. Consequently,algae response to heavy metal stress at multiple levels,more records with different angle and levels are thus still required to evaluate their effects on algae in the future.
杂志排行
Journal of Oceanology and Limnology的其它文章
- Response of the North Pacific Oscillation to global warming in the models of the Intergovernmental Panel on Climate Change Fourth Assessment Report*
- Effect of mesoscale wind stress-SST coupling on the Kuroshio extension jet*
- Surface diurnal warming in the East China Sea derived from satellite remote sensing*
- Cross-shelf transport induced by coastal trapped waves along the coast of East China Sea*
- Observations of near-inertial waves induced by parametric subharmonic instability*
- Seasonal variation and modal content ofinternal tides in the northern South China Sea*