不同冠层结构下的植物生长型与生活型特征
2018-07-04张星元蔡琳颖
张星元,张 璐,马 丁,蔡琳颖
(华南农业大学 林学与风景园林学院,广东 广州 510642)
森林冠层是影响森林生态系统特性最基本的因素[1],是森林生态系统与外界环境作用最直接与最活跃的界面[2]。我国对森林冠层结构的研究起步较晚,随着林冠实测技术的发展,冠层研究从定性描述发展为更加严格的量化研究[3-4]。而国外学者对森林冠层结构、林下植被及光环境特征的研究已有30多年,目前已有大量的研究成果[5-6]。林木冠层通过对光照进行吸收、反射和投射,改变林下光照条件,从而对林下植被的物种组成与物种多度产生影响[7-8]。根据冠层结构在森林群落中位置的不同,可将其分为林下、林隙边缘和林隙中心。研究森林群落不同冠层结构下植物生长型和生活型的特征,不仅可为森林生态系统小气候、群落演替及其进化史等提供关键信息[9],也可加强对整个森林生态系统演替进程及其机制的理解[10]。
植物生长型和生活型深刻解析植物群落结构及其与环境的关系,对研究植物群落的发生、发展以及演替规律具有重要意义[11]。植物的生活型是植物对于综合生境条件长期适应而在外貌上反映出来的植物类型[12]。森林群落的外貌主要取决于生活型的组成,通过对植物群落生活型的研究既可以发现控制和影响群落的主要气候因素、植物群落与环境之间的关系,又可以了解群落组成种的外貌特征随着地理位置或生境的改变而发生的变化[13]。植物生长型是指根据植物的可见结构划分的类群,是表征群落外貌特征和垂直结构的重要指标[14]。植物的生长型反映植物生活的环境条件,相同的环境条件具有相似的生长型,是趋同适应的结果[15]。
该研究以南岭山地森林群落为研究对象,量化其冠层结构、光环境特性与林下植被生长型和生活型的关系,以期为亚热带森林群落更新和维持提供理论和数据支持。主要研究以下问题:南岭山地森林林下植被在林下、林隙边缘和林隙中心这3种冠层结构下的生长型和生活型表现特征是否都遵循同一模式?该地区森林群落的物种分布对不同冠层结构参数的响应是否一致?
1 材料与方法
1.1 研究区概况
研究试验地设在广东南岭国家级自然保护区内(112°41′~ 113°15′E,24°39′~ 28°08′N),总面积58 368.4 hm2。保护区位于广东北部南岭山脉南坡中段,是广东省陆地森林面积最大的自然保护区,保存有较完整的中亚热带针叶林、针阔混交林、常绿阔叶林、山地矮林等[16]。该区位于中亚热带湿润性季风气候区,海拔500~600 m,年均气温17.4 ℃ ,最冷月(1月份)平均气温7.1 ℃ ,最热月(7月份)平均气温26.2 ℃ ,年均降雨量2 108.4 mm,年平均相对湿度84%,日照率40%。水平地带性土壤为红壤,地带性植被为亚热带常绿阔叶林,森林覆盖率达98%[17]。
1.2 样方设置与调查
在南岭国家级自然保护区,以亚热带山地森林群落为研究对象,采用样方法,以10 m×10 m为样方单元,在每个样方单元内进行每木调查,测定胸径(DBH)≥ 3 cm的所有活立木的种名、胸径、树高和冠幅,共计调查了100个样方(10 000 m2)。同时,在每个样方单元的四角和对角线中心各设置1个2 m× 2 m的小样方,记录每个小样方内林下植物的种名、多度与盖度。
1.3 半球面影像拍摄与分析
半球面影像技术是一种快捷、准确、高性价比的近距离遥感方法,近年来成功应用于森林生态监测和评价研究中。具体方法为:用三脚架将Nikon CoolPix 4500 数码相机外接Nikkor FC-E8 鱼眼镜头转换器水平放置于离地面1.65 m处,用指南针确定方向使记录的照片顶部与磁北方向重合,在每个10 m × 10 m样方单元中心和对角线四分位处镜头朝上拍摄半球面林冠影像。采用Gap Light Analyzer 2.0 (GLA)图像处理软件分析林冠影像,输出林冠开度(CO)、叶面积指数(LAI)、林下直射光(Transmitted direct light, Tdir)、散射光(Transmitted diffused light, Tdif )和总光照(Transmitted total light, Ttot) 5个冠层结构参数,并进行冠层结构参数分组[18-20]。
1.4 数据分析
参照Whittaker[21]的群落分类系统,结合亚热带山地森林的植物类型,将森林群落维管束植物按生长型分为乔木、灌木、草本、藤本和蕨类五个层次。参照Raunkiaer[22]的分类系统,将群落按生活型分为地面芽、地上芽、高位芽3个类别。高位芽又进一步分为藤本高位芽、矮高位芽、小高位芽和中高位芽。同时,参照樊大勇等[23]的林冠划分依据,并结合实地观测,将林冠开度从0~7%定为林下,7%~14%定为林隙边缘,19%~21%定为林隙中心。运用Statistica 8.0统计软件对不同冠层结构下植被生长型和生活型的物种多度以及冠层的叶面积指数、林冠开度和林下散射光等5个结构参数分别进行Kruskal-Wallis(非参数ANOVA)分析,检验物种多度在各分组变量间是否存在差异。运用PC-ORD 6.0多元分析软件对山地森林群落冠层结构各参数典范对应分析(Canonical correspondence analysis, CCA),并采用蒙特卡罗检验来检验结果是否具有统计显著性。
2 结果与分析
2.1 不同冠层结构下的植物生长型和生活型
2.1.1 不同冠层结构下的植物生长型特征
在不同冠层结构下,森林群落乔木多度均显著大于草本、藤本和蕨类的多度,说明乔木在南岭山地森林不同冠层结构下均占据优势地位。灌木的优势地位也较明显,仅次于乔木,而草本、藤本以及蕨类的丰富度与多度表现不尽一致(见图1)。其中,林下草本与藤本的丰富度和多度均小于其在林隙边缘和林隙中心,蕨类在林隙边缘的多度最大,说明草本与藤本在林下的生长优势最弱,而蕨类最适于在林隙边缘生长。
2.1.2 不同冠层结构下的植物生活型特征
基于10 000 m2样地的调查数据,共记录到高位芽植物297种、地面芽植物36种、地上芽植物13种。在不同冠层结构下,植物群落丰富度与多度的生活型数量特征表现基本一致(见图2),表明研究区植物群落的生活环境适宜而稳定。随着林冠开度的增加,地面芽、地上芽和高位芽植物的物种丰富度和多度均表现为先增后减,说明在该地区林隙边缘的生活环境更适合植物群落的生长。对群落高位芽植物进一步的分析得知,在林下、林隙边缘和林隙中心这3种不同的冠层结构下,各高位芽植物的丰富度与多度的特征表现一致,从小到大依次为藤本高位芽,矮高位芽,小高位芽,中高位芽。其中,中高位芽植物优势明显,而地上芽植物最少,在林下这一冠层结构下没有地上芽植物,藤本高位芽也较少,表明在中亚热带山地森林适宜中高位芽的生长,不适宜地上芽和藤本高位芽的生长。
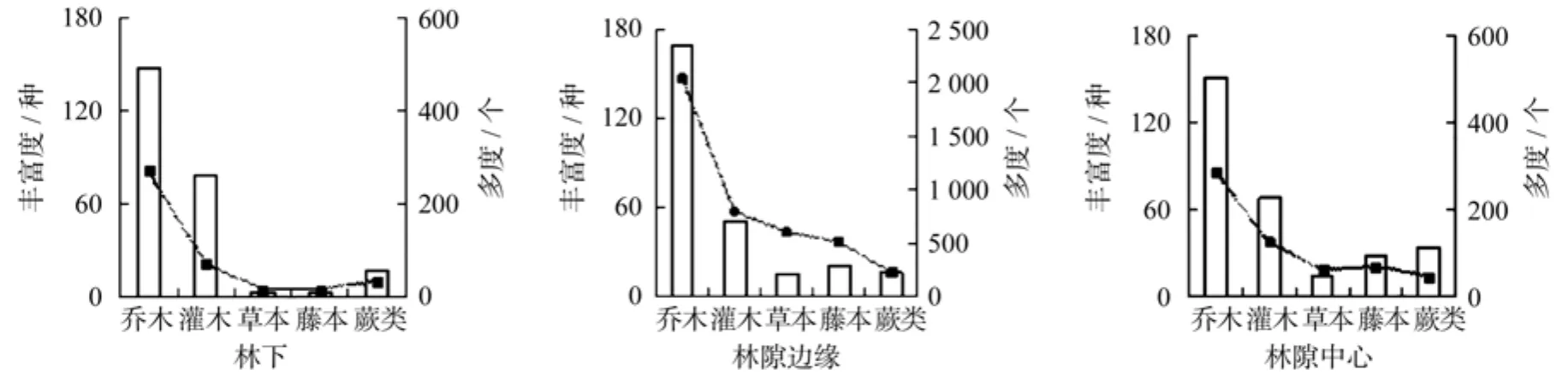
图1 不同冠层结构下的植物生长型数量特征Fig.1 Plant growth forms characteristics of different canopy structures in subtropical mountainous forest
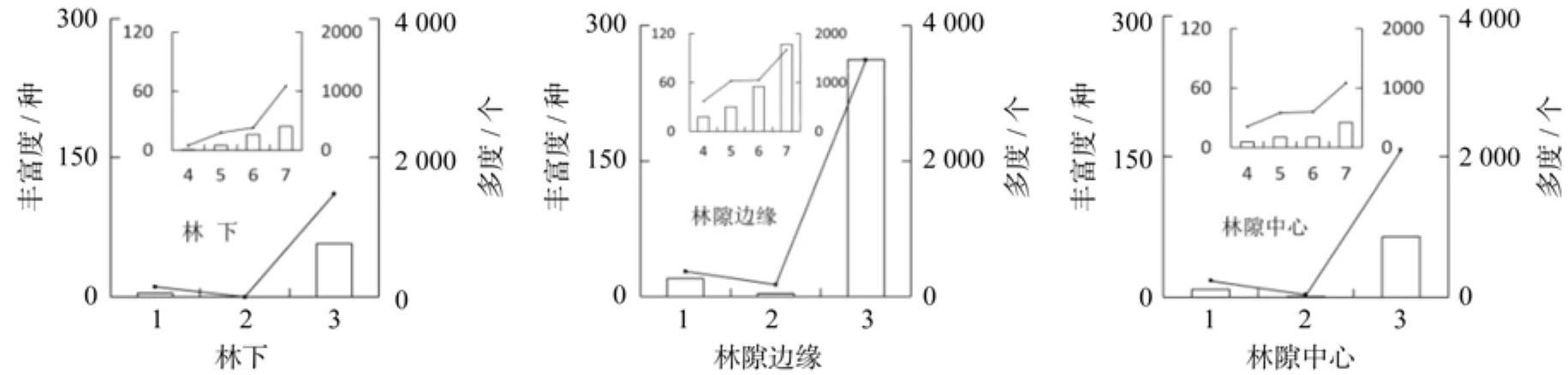
图2 不同冠层结构下的植物生活型数量特征Fig.2 Characteristics of plant life forms of different canopy structures in subtropical mountainous forest
2.2 不同冠层结构下植物生长型和生活型的差异性分析
对不同冠层结构下各个生长型的多度进行的Kruskal-Wallis分析表明(见图3),灌木多度呈极显著差异(P<0.001),草本植物表现为显著差异(P<0.05),但乔木、藤本与蕨类植物的差异性水平并不显著(P>0.05)。多重比较进一步表明,林下的草本多度与其他2个冠层结构差异显著,说明林下的生活环境不利于草本植物的生长;林下和林隙边缘的灌木多度差异显著,但林隙中心的灌木多度与林下和林隙边缘都没有达到显著性水平。
与生长型类似,3个冠层结构下的中高位芽、藤本高位芽与地面芽植物多度的差异性水平也不显著(P>0.05)(见图4)。多重比较进一步发现,小高位芽在林下和林隙中心这2种冠层结构下差异显著(P<0.05),但在林隙边缘差异不显著(P>0.05)。矮高位芽只在林隙中心与林下和林隙边缘存在显著差异(P<0.05)。

图3 不同冠层结构下的植物生长型差异性分析Fig.3 Kruskal-Wallis analysis of growth forms of different canopy structures in subtropical mountainous forest
2.3 不同生长型和生活型植物分布对冠层结构的响应

图4 不同冠层结构下的植物生活型差异性分析Fig.4 Kruskal-Wallis analysis of life forms of different canopy structures in subtropical mountainous forest
冠层结构参数与环境轴的相关系数以及不同冠层结构下的CCA特征根如表1所示,在林下,CO和LAI与环境轴2相关性最大,而Tdir、Tdif和Ttot与环境轴1相关性最大;在林隙边缘,CO、LAI、Tdif和Ttot均与环境轴3最具相关性,而Tdir与环境轴1相关性最大;在林隙中心,五种冠层结构参数均与环境轴1最具相关性。同时,蒙特卡罗检验特征值以及物种与环境因子的相关系数P 值皆小于0.05,说明排序效果理想。
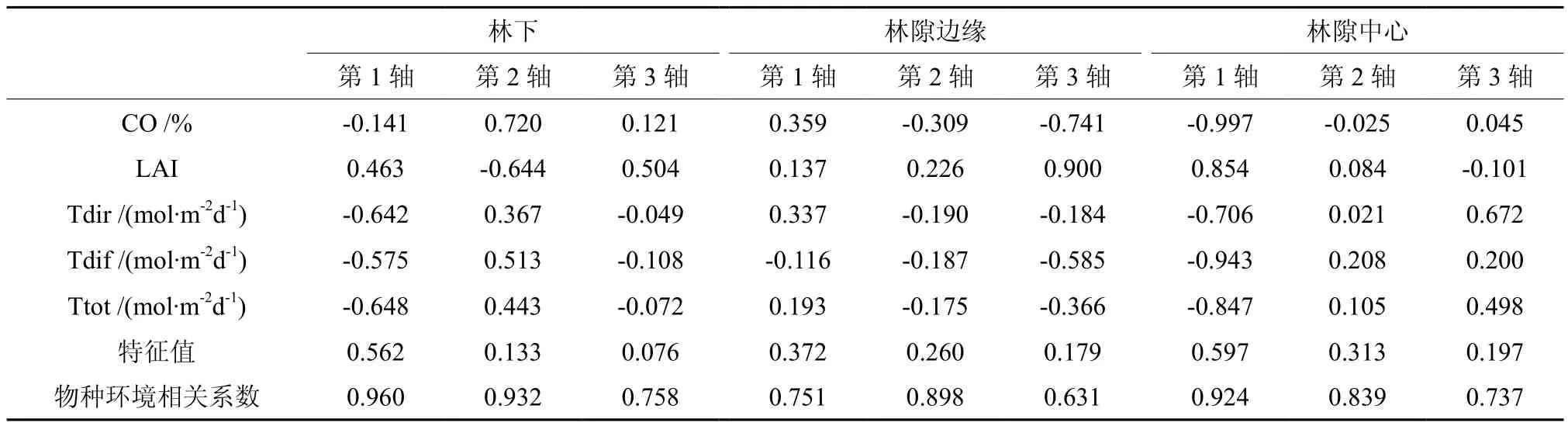
表1 不同冠层结构下各参数与环境轴的相关系数以及特征根Table 1 The correlation coefficient and characteristic roots of parameters and environmental axis under different canopy structures
CCA排序结果显示(见图5和表2),在林下和林隙中心,不同生长型和生活型植物受到5个冠层结构参数的影响,而在林隙边缘,物种分布只与叶面积指数、林冠开度和林下散射光有关。在林下,腺萼马银花Rhododendron bachii、杨桐Adinandra millettii和黄丹木姜子Litsea elongata等集中分布在CCA排序图中林冠开度较小或叶面积指数较大的区域,表明这些物种较喜欢阴蔽的环境或具备耐阴性。在林隙边缘,华山姜Alpinia oblongifolia、石斑木Raphiolepis indica和乐东拟单性木兰Parakmeria lotungensis等分布在林冠开度较大的区域,表明这些物种适合生长在林冠光照较强的区域。在林隙中心,假地枫皮Illicium jiadifengpi和青冈Cyclobalanopsis glauca等分布在林冠开度较大的区域,而檵木Loropetalum chinense分布在林下直射光较大的区域,说明这几种植物都具喜阳特性。藤本植物山蒟Piper hancei、龙须藤Bauhinia championii和瓜馥木Fissistigma oldhamii等均分布在光照参数周围,受光照影响较大,且藤本植物对林下直射光的依赖性较强。在林隙中心的大多数蕨类,如阔鳞毛蕨Dryopteris championii、单叶新月蕨Pronephrium simplex和深绿卷柏Selaginella doederleinii等均分布在叶面积指数较大的区域,说明多数蕨类植物尽管生长在林冠开度较大的环境中,却仍属耐阴性植物。

图5 不同冠层结构下的优势种与环境因子的CCA分析Fig.5 CCA analysis of dominant tree species and environmental factors in different canopy strutures in subtropical mountainous forest

表2 CCA排序图物种名称及代码Table 2 The species and species code of CCA

续表2Continuation of table 2
3 结论与讨论
3.1 森林群落冠层结构与植物生长型和生活型的关系
相对于密集的冠层结构,草本植物在稀疏冠层结构下的生物量会更多[24]。在林隙光照强的地方,林下植物的多度相对较高[25]。Gálhidy等[26]认为草本植物在大林隙中的平均覆盖度和丰富度均比小林隙中的大。王永强等[27]认为,在林冠开度为10%~30%时,林下植物的多样性随林冠开度的增加略有增加,表现为不显著相关。该研究表明,不同冠层结构下乔木和灌木的多度均显著大于草本、藤本和蕨类。其中,乔木、藤本和蕨类植物没有显著性差异,而灌木和草本则差异显著,说明冠层结构对灌木和草本植物的生长影响较大。同时,不同冠层结构下,矮高位芽、小高位芽和地上芽均表现为显著差异,说明不同的冠层结构对植物的生活型有不同的影响。而地上芽在林下的多度为零,可能是由于地上芽对光有较大需求,不适合在郁闭度较大的林下生长。
3.2 森林群落冠层结构与物种分布的关系
冠层通过影响林下小气候、改变植被生长条件[28],从而对林下植被的生长、更新与物种组成产生影响[29-30]。该研究表明,不同冠层结构下植物的物种组成与分布表现不一。林冠开度与林下光照条件直接相关,可通过影响林下光照进而影响植被生长[17]。其中,红楠Machilus thunbergii和少叶黄杞Engelhardtia fenzelii等均分布在林冠开度较小的阴蔽环境,这与房震等[31]研究中指出红楠喜湿耐阴林隙边缘的结论一致。而华山姜、石斑木和乐东拟单性木兰等物种则分布在林冠开度较大的区域。山蒟、龙须藤和瓜馥木等藤本高位芽植物均分布在直射光较大的强光区,阔鳞毛蕨、单叶新月蕨和深绿卷柏等蕨类植物则分布在叶面积指数较大的区域,说明藤本植物比蕨类植物更具喜光性。毛果柃Eurya trichocarpaKorth、箬竹Indocalamus tessellatus、粘毛杜鹃Rhododendron championae与黄丹木姜子这四种植物在不同冠层结构下均有分布,表明这4种植物具较强适应性并在群落中占据优势地位。同时,冠层结构参数与植物分布也具相关关系,林冠开度(CO)和叶面积指数(LAI)均与环境轴的夹角较小,表明与植物分布的相关性较强,且CO和光照因子均与LAI成负相关,这与崔佳玉等[32]的研究一致。
研究中发现,一些珍稀濒危植物如金毛狗Cibotium barometz、白豆杉Pseudotaxus chienii和白桂木Artocarpus hypargyreus等仅生长于林隙边缘或林隙中心,乐东拟单性木兰、华南五针松Pinus kwangtungensis和福建柏Fokienia hodginsii等仅在林下和林隙边缘有分布。还有一些具有观赏价值或者药用价值的植物如檵木Loropetalum chinense和假地枫皮Illicium jiadifengpi等在林隙中心生长较好,戟叶圣蕨Dictyocline sagittifolia和光里白Diplopterygium laevissima等则分布在林下叶面积指数较大的区域。探究各冠层结构下不同功能性植物的分布特征,可明确其生长习性,并采取适当保护措施。
该研究还表明,林隙边缘的生活环境更适合植物群落的生长。相对于林冠开度较小的林下和林冠开度较大的林隙中心,林冠开度中等的林隙边缘的地面芽、地上芽和高位芽植物的物种丰富度和多度均较大。这与Connell[33]的“中等干扰假说”相符,即物种丰富度在受到中等强度干扰的环境中最大,在未受干扰和受到高强度干扰的环境中则相对较小。由于在未受干扰时,存在大量的先锋种,不利于其它物种的生长,有的甚至被排挤掉;相反,在受高强度干扰的环境中,大量物种因无法承受干扰而消失,只有少量的耐受性较高的物种还存在;而在中等强度干扰的环境中,先锋种受到一定的限制,同时又不至使一些物种因干扰而消失,因此大量的物种可以共存,物种丰富度最高。臧润国等[34]研究的南亚热带常绿阔叶林不同大小和发育阶段林隙的树种多样性,结果也显示物种丰富度和多样性指数随林隙大小的变化呈现单峰型趋势。
该研究采用间接梯度分析的方法,分析各物种的生活型和生长型特征及其与林下不同生境的关系,探讨了山地森林中优势种对光环境的选择,明确其耐阴或喜光特性,不但为研究亚热带山地森林生态系统更新和维持其生物多样性提供理论和数据支持,还可为城市园林及药用植物物种驯化和植物选择提供科学参考,并为亚热带森林生态系统多样性保护和珍稀濒危物种保育提供科学依据。但由于森林冠层结构的复杂性,对于林冠结构的形成过程、林冠成熟及达到顶级时期的动态变化和不同时期冠层结构形成的光环境条件对林下植被的影响,还没有做到很系统的研究。而叶面积指数和林冠开度应该可以指示这种时间和空间上的动态变化,但要确定其关系,还有待进一步的研究。
[1]Hansen A J, Phillips L B, Dubayah R,et al.Regional-scale application of lidar: Variation in forest canopy structure across the southeastern US[J].Forest Ecology and Management,2014,329: 214-226.
[2]敬小丽,张 璐,杜伟静,等.南岭山地森林群落冠层结构对林下野生花卉的影响[J].西南农业学报,2015,28(2):833-838.
[3]任 海,彭少麟.鼎湖山森林群落的几种叶面积指数测定方法的比较[J].生态学报,1997, 17(2):110-113.
[4]李德志,臧润国.森林冠层结构与功能及其时空变化研究进展[J].世界林业研究,2004, 17(3):12-16.
[5]Ediriweera S, Singhakumara B M P, Ashton M S.Variation in canopy structure, light and soil nutrition across elevation of a Sri Lankan tropical rain forest[J].Forest Ecology and Management,2008, 256(6): 1339-1349.
[6]Gillespie T W, de Goede J, Aguilar L,et al.Predicting tree species richness in urban forests[J].Urban Ecosystems,2017,20(4): 839-849.
[7]Brunet J R, Valtinat K, Mayr M L,et al.Understory succession in post-agricultural oak forests: Habitat fragmentation affects forest specialists and generalists differently[J].Forest Ecology and Management.2011, 262(9): 1863-1871.
[8]Yu M, Sun O J.Effects of forest patch type and site on herb-layer vegetation in a temperate forest ecosystem[J].Forest Ecology and Management,2013, 300(300): 14-20.
[9]Leibold J M C A.Ecological Niches: Linking Classical and Contemporary Approaches[J].Biodiversity and Conservation,2004, 13(9): 1791-1793.
[10]Steenberg J W N, Millward A A, Nowak D J,et al.Forecasting Urban Forest Ecosystem Structure, Function, and Vulnerability[J].Environmental Management,2017,59(3):373-392.
[11]侯志勇,谢永宏,陈心胜,等.洞庭湖湿地植物生活型与生态型[J].湖泊科学,2016,28(5): 1095-1102.
[12]黎燕琼,郑绍伟,龚固堂,等.米仓山巴山水青冈群落结构特征及更新状况分析[J].四川林业科技,2014,35(2):44-46.
[13]薛 睿,张继强,王三英,等.甘肃陇南地震扰动区植被群落特征[J].草业科学,2015,32(11): 1774-1780.
[14]Rowe N S T.Plant growth forms: An ecological and evolutionary perspective[J].New Phytologist,2005, 166(1): 61-72.
[15]周会萍,职丽娟,蔡祖国,等.江西吉安退化湿地松群落外貌特征分析[J].江西农业大学学报,2009,31(4):690-694.
[16]程 真,周光益,吴仲民,等.南岭南坡中段不同群落林下幼树的生物多样性及分布[J].林业科学研究,2015,28(4):543-550.
[17]敬小丽,杜伟静,张 璐,等.南岭山地森林群落冠层结构与立木多度的关系[J].中南林业科技大学学报,2014,34(5):59-65.
[18]Soudani K, Trautmann J,et al.Leaf area index and canopy stratification in Scots pine (Pinus sylvestrisL.) stands[J].International Journal of Remote Sensing,2002,23(18):3605-3618.
[19]Lertzman KP, Sutherland GD, Inselberg A,et al.Canopy gaps and the landscape mosaic in a coastal temperate rain forest[J].Ecology,1996,77(4):1254-1270.
[20]苏志尧,贾小容,解丹丹,等.用半球面影像技术测定城市林分的冠层结构参数[J].华南农业大学学报,2007,28(2):65-68.
[21]Whittaker R H.Communities and ecosystems(2nd Revised edition)[M].Prentice Hall (a Pearson Education Company): New Jersey, 1975:1-352.
[22]Raunkiaer C.The life-forms of plants and statistical geography[M].Oxford:The Clarendon Press: 1934:1-359.
[23]樊大勇,张旺峰,陈志刚,等.沿林冠开度梯度的银杉幼树对光的适应性[J].植物生态学报, 2005,29(5):713-723.
[24]Shirima D D, Pfeifer M, Platts P J,et al.Interactions between Canopy Structure and Herbaceous Biomass along Environmental Gradients in Moist Forest and Dry Miombo Woodland of Tanzania[J].PLOS ONE,2015,10(11): e142784.
[25]孙 东,朱 纯,熊咏梅,等.林隙光照对广州市风景林林下植物分布的影响[J].华南农业大学学报,2011,32(4):63-66.
[26]Lhidy L S, Mih K B, Hagy A,et al.Effects of gap size and associated changes in light and soil moisture on the understorey vegetation of a Hungarian beech forest[J].Plant Ecology, 2006,183(1): 133-145.
[27]王永强,蔡燕茹,曾焕忱,等.不同林冠开度下亚热带林下植物的组成和多样性[J].西北农林科技大学学报(自然科学版),2016,44(5):64-72.
[28]Dumais D, Prévost M.Physiology and growth of advance Picea rubens and Abies balsamea regeneration following different canopy openings[J].Tree Physiology,2014,34(2):194-204.
[29]Mori A S, Fukasawa Y, Takeda H.Tree mortality and habitat shifts in the regeneration trajectory underneath canopy of an oldgrowth subalpine forest[J].Forest Ecology and Management,2008, 255(11): 3758-3767.
[30]Vizoso-Arribe O, Díaz-Maroto I, Vila-Lameiro P,et al.Influence of the canopy in the natural regeneration of Quercus robur in NW Spain[J].Biologia,2014,69(12):1678-1684.
[31]房 震.红楠的特征特性及育苗技术[J].现代农业科技,2012(6):220-221.
[32]崔佳玉,曾焕忱,王永强,等.银瓶山自然保护区阔叶林冠层结构与辐射消减效应[J].西北林学院学报,2015,30(4):45-49.
[33]Connell J H.Diversity in tropical rain forest and coral reefs[J].Science,1978,199: 1302-1310.
[34]臧润国,王伯荪,刘静艳.南亚热带常绿阔叶林不同大小和发育阶段林隙的树种多样性研究[J].应用生态学报,2000,11(4):485-488.
