低温胁迫对不同小麦品种结实率和活性氧代谢的影响
2018-06-29陈巧艳李迎迎陈刘平欧行奇赵星星张自阳刘明久朱启迪
陈巧艳, 李迎迎, 陈刘平, 欧行奇, 赵星星, 张自阳, 刘明久, 朱启迪
(河南科技学院,河南新乡 453003)
在人类的生活中,小麦是主要的粮食作物之一。小麦在全球粮食作物中占有不可或缺的地位,全球大约有35%以上的人口以小麦为主要口粮[1]。我国的第一大麦区为黄淮南片麦区,河南省小麦产区属于黄淮南片麦区,其面积和总产量都占全国的40%以上。黄淮麦区经常发生霜冻,是我国冬小麦霜冻害的高发区,自20世纪80年代以来,黄淮麦区发生霜冻害的频率达到45%,特别是商丘地区已高达60%[2]。2004年,黄淮麦区发生大面积的早春积霜冻害,受害冬小麦面积为333 hm2[3]。近些年来,随着全球气候变暖,无霜期的时间缩短,干旱越来越严重,这种变化使得春季的气候变得不稳定,使得春冻出现的频率增多,在黄淮南片麦区春季霜冻对小麦的生产已经产生严重影响,制约着该产区小麦的高产稳产,已经成为该地区的主要农业气象灾害之一[4-5],因此研究低温胁迫对小麦生理生化反应的影响至关重要。本研究以半冬性小麦品种百农207、AK58、郑麦366为材料,研究低温胁迫对小麦结实率、抗氧化酶活性以及渗透调节物质含量的影响,阐述小麦对低温胁迫的响应机制。
1 材料与方法
1.1 试验材料
选用3个半冬性小麦品种百农207、AK58、郑麦366,均由河南科技学院百农207团队提供。
1.2 试验方法
本试验于2016—2017年在河南省新乡市河南科技学院作物育种实验室进行,每品种挑选大小一致、籽粒饱满的种子,先用70%乙醇活化30 s,再用0.1% HgCl2消毒10 min,然后用自来水冲洗5次,无离子水冲洗3次。将消毒好的种子种在盆里,每盆20粒种子,然后放入人工气候室中培养。待小麦幼穗发育到雌雄蕊原基分化期时,放入人工气候箱于0 ℃低温处理3 d,对照于正常环境中生长,取小麦叶片放入 -80 ℃ 低温冰箱备用。
1.3 测定指标与方法
叶绿素鲜质量含量测定采用丙酮乙醇混合浸提法[6];脯氨酸含量测定采用茚三酮法[7];过氧化物酶(POD)活性(鲜质量)测定采用愈创木酚法[8];超氧化物歧化酶(SOD)活性(鲜质量)测定采用氮蓝四唑法[8];丙二醛(MDA)含量采用硫代巴比妥酸法测定[9];在小麦扬花期前,在正常生长条件和低温处理条件下各随机挑选10个小麦幼穗进行人工套袋,成熟期调查小麦的结实性。
1.4 数据处理
采用Excel 2010和DPS软件对试验数据进行统计分析,采用Duncan氏新复极差法进行多重比较。
2 结果与分析
2.1 低温胁迫对不同小麦品种结实率的影响
由表1可以看出,在正常生长情况下,AK58的结实率较高,达到99.3%,其次是百农207、郑麦366;在低温处理条件下,AK58、百农207的结实率依然较高,但是郑麦366受低温的影响,结实率骤然下降,仅为9.7%。
2.2 低温胁迫对不同小麦品种叶绿素含量的影响
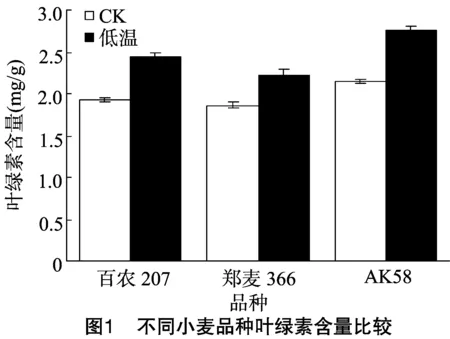
从图1可以看出,低温下3个品种叶绿素含量均比对照增高,百农207、郑麦366、AK58分别较对照增加了27.8%、19.9%、28.1%。 百农207、 AK58 的叶绿素含量在低温处理与对照之间差异较大,而郑麦366差异较小。

表1 不同小麦品种结实率的比较
注:同列数据后不同小写字母表示同一处理不同品种间差异达0.05显著水平。
2.3 低温胁迫对不同小麦品种SOD活性的影响
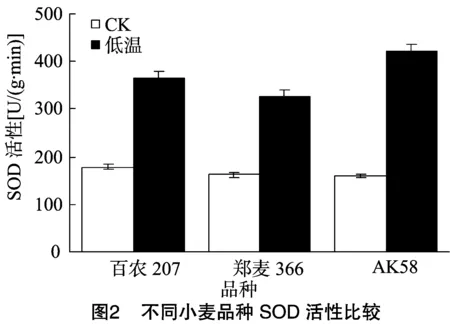
SOD是膜脂过氧化防御系统的主要保护酶,能催化活性氧发生歧化反应,产生无毒O2和H2O2,从而避免植物遭受伤害。由图2可知,在正常生长条件下,百农207、郑麦366、AK58的SOD活性分别为178.6、162.7、159.3 U/(g·min);低温胁迫后,百农207、郑麦366、AK58 3个品种SOD活性均上升,其中百农207和AK58受到低温胁迫后其SOD活性较高。
2.4 低温胁迫对不同小麦品种POD活性的影响
由图3可知,低温胁迫下,百农207、郑麦366、AK58的POD活性均比对照增加,其中百农207、AK58的POD活性上升幅度高于郑麦366,表明百农207、AK58受到低温胁迫后其POD活性较高,更有利于清除过氧化物,降低活性氧对细胞的损伤。
2.5 低温胁迫对不同小麦品种MDA含量的影响
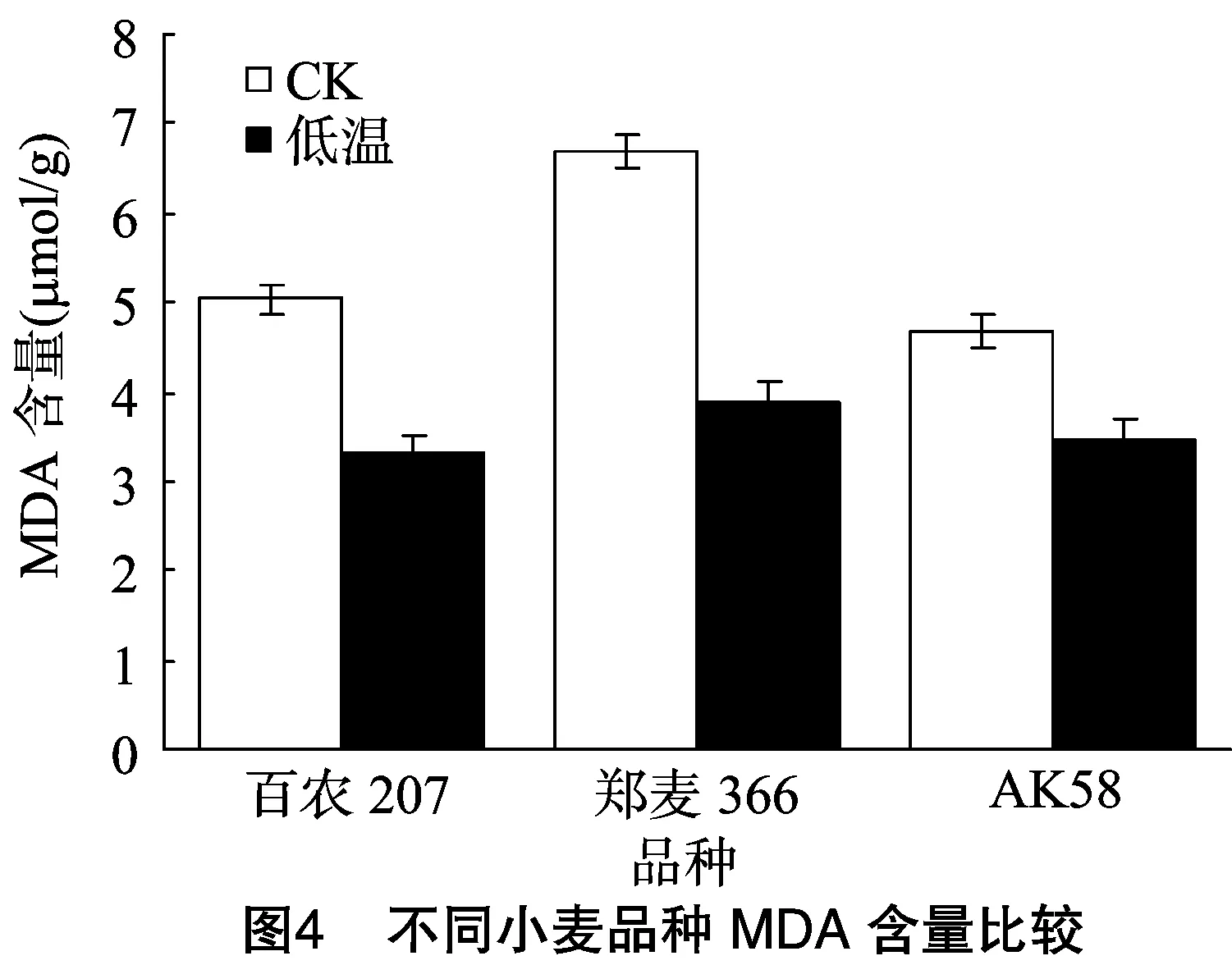
丙二醛是膜脂过氧化终产物,是膜系统受伤害程度的重要标志之一,其含量越高,细胞膜系统受到逆境胁迫的伤害越严重。由图4可知,在正常条件下,郑麦366丙二醛含量较高,而百农207和AK58丙二醛含量较低;低温处理后,3个品种的丙二醛含量均下降,郑麦366丙二醛含量仍然高于百农207和AK58。

2.6 低温胁迫对不同小麦品种脯氨酸含量的影响
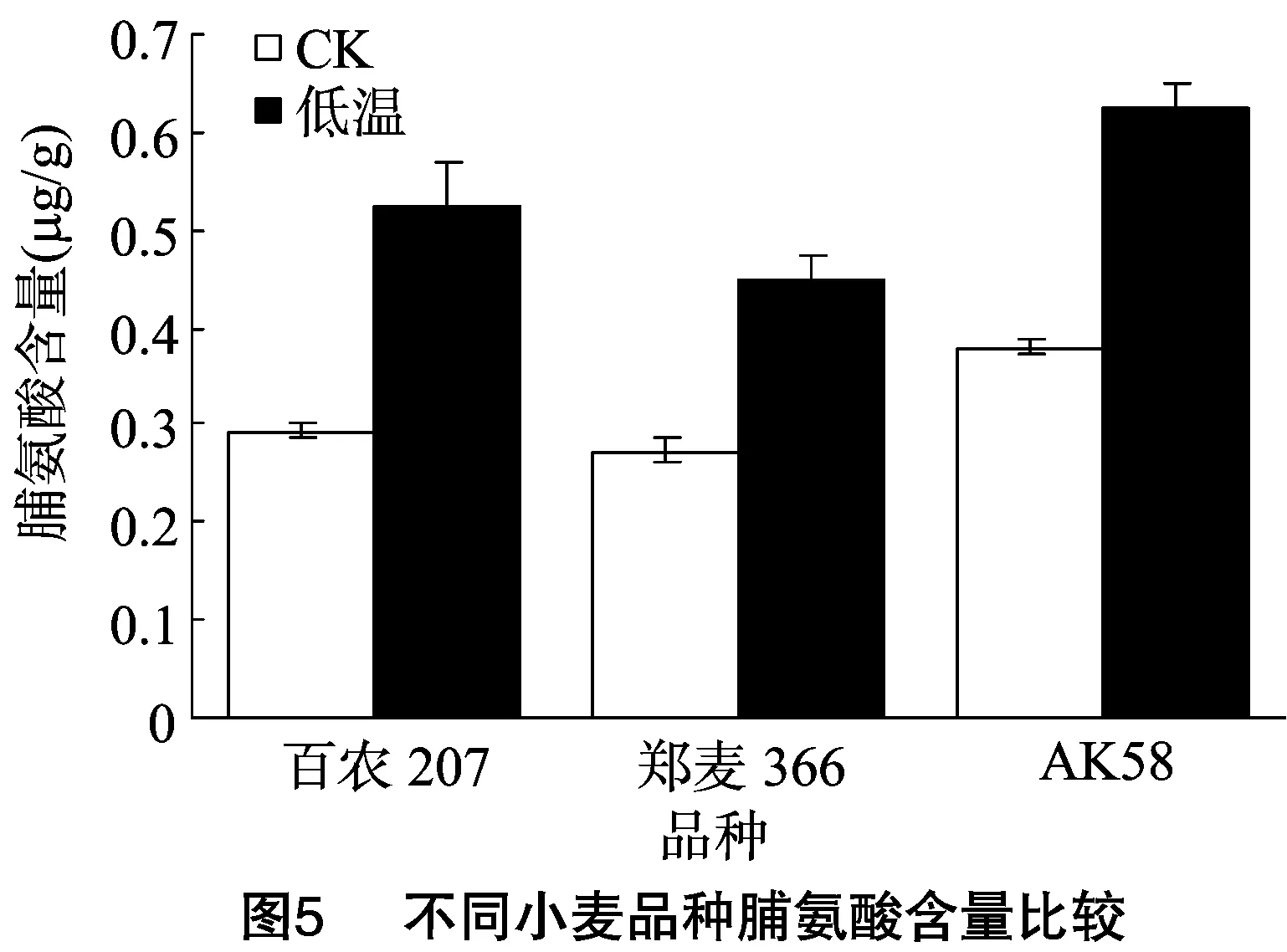
由图5可知,在正常条件下,百农207、AK58的脯氨酸含量较高;低温胁迫后,3个品种脯氨酸含量均增加,其中AK58、百农207脯氨酸含量增加得较多,而郑麦366脯氨酸含量增加得较少,表明受到低温胁迫后,AK58、百农207脯氨酸含量较高,具有较强的细胞渗透调节能力。
3 讨论与结论
有研究表明,倒春寒严重影响小麦的结实性,使小麦的结实率下降,最终影响小麦的产量。本研究发现,在低温胁迫下,百农207、AK58的结实率降低不明显,而郑麦366下降比较明显,表明百农207、AK58抗倒春寒能力强,而郑麦366的抗倒春寒能力较弱。众多研究者认为,低温胁迫使叶绿素含量下降。在低温环境中,植物叶片细胞内活性氧大量积累,抑制叶绿体色素生物合成酶的活性[10],阻碍叶绿素合成并导致叶绿素降解。本试验发现,在低温处理3 d后,3个品种叶绿素含量均表现为上升。可能是植物对低温逆境的应激保护反应,通过加快叶绿素合成来补偿和提高叶片对光能的吸收、传递、转换能力,合成速度大于降解速度,使叶绿素含量升高。
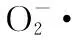
有学者研宄表明,低温胁迫导致可溶性物质积累[20-21],既降低细胞中的水势,提高细胞的保水能力,又可以保护生物大分子,防止其失活,例如低温诱导蛋白可以提高植物抗冷性的作用[22]。脯氨酸被认为是重要的渗透调节物质,不仅可以维持细胞渗透调节能力,阻止水分丧失,还是可溶性溶质中唯一可整合单线态氧、高效清除羟自由基的分子,有助于稳定蛋白质和膜系统,还能作为含氮的储藏物质和恢复生长的能源,提高植物抗冻性[23]。本研究发现,低温胁迫引起脯氨酸含量上升,这与他人的研究结果[24-27]一致。与郑麦366相比,受到低温胁迫后百农207、AK58的脯氨酸含量较高,表明在低温胁迫条件下,百农207、AK58的脯氨酸含量较高,能够提高细胞的渗透调节能力,进而提高抗寒能力。
参考文献:
[1]倪胜利,张国宏,李兴茂. 小麦抗寒性研究概述[J]. 甘肃农业科技,2008(8):23-26.
[2]赵苗稳,尚红敏,司马青焕,等. 2010年冬小麦晚霜冻害调查及成因分析[J]. 农村经济与科技,2010,21(9):107-120.
[3]高 媛,齐晓花,杨景华,等. 高等植物对低温胁迫的响应研究[J]. 北方园艺,2007(10):58-61.
[4]余松烈. 中国小麦栽培理论与实践[M]. 上海:上海科学技术出版社,2006:65.
[5]王绍中,郑天存,郭天财. 河南小麦育种栽培研究进展[M]. 北京:中国农业科学技术出版社,2007.
[6]张宪政. 作物生理研究法[M]. 北京:农业出版社,1992:129-150.
[7]高俊风. 植物生理学实验技术[M]. 北京:世界图书出版社,2000:192-193.
[8]孙 群,胡景江. 植物生理研究技术[M]. 杨凌:西北农林科技大学出版社,2005:176-177.
[9]赵海泉. 基础生物学实验指导-植物生理学分册[M]. 北京:中国农业大学出版社,2008:65-81.
[10]苗 微,王国骄,马殿荣,等. 辽宁省杂草稻幼苗对低温胁迫的生理响应[J]. 中国水稻科学,2011,25(6):639-644.
[11]Baek K H,Skinner D Z. Alteration of antioxidant enzyme gene expression during cold acclimation of near-isogenic wheat lines[J]. Plant Science,2003,165(6):1221-1227.
[12]Xu S,Li J L,Zhang X Q,et al. Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation,antioxidant metabolites and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress[J]. Environmental and Experimental Botany,2006,56(3):274-285.
[13]Saruyama H,Tanida M. Effect of chilling on activated oxygen scavenging enzymes in low temperature-sensitive and tolerant cultivars of rice[J]. Plant Science,1995,109(2):105-113.
[14]刘慧英,朱祝军,吕国华. 低温胁迫对嫁接西瓜耐冷性和活性氧清除系统的影响[J]. 应用生态学报,2004,15(4):659-662.
[15]李品明,孙玉芳,杨丙贤,等. 低温胁迫对黄连膜脂过氧化作用和抗氧化酶活性的影响[J]. 中国农学通报,2011,27(15):117-120.
[16]张俊环,黄卫东. 葡萄幼苗在温度逆境交叉适应过程中活性氧及抗氧化酶的变化[J]. 园艺学报,2007,34(5):1073-1080.
[17]刘 伟,艾希珍,梁文娟,等. 低温弱光下水杨酸对黄瓜幼苗光合作用及抗氧化酶活性的影响[J]. 应用生态学报,2009,20(2):441-445.
[18]高小丽,孙健敏,高金峰,等. 不同基因型绿豆叶片衰老与活性氧代谢研究[J]. 中国农业科学,2008,41(9):2873-2880.
[19]罗 娅,汤浩茹. “丰香”草莓果实发育过程中抗氧化物质与活性氧代谢研究[J]. 园艺学报,2011,38(8):1523-1530.
[20]齐阳阳,杜晓华,王梦叶,等. 大花三色堇和角堇对低温胁迫的生理响应及其抗寒性[J]. 江苏农业科学,2017,45(15):115-118.
[21]刘彤彤,蒋欣梅,于锡宏,等. 间歇降温对黄瓜幼苗耐冷相关指标的影响[J]. 江苏农业科学,2016,44(12):195-196,271.
[22]薛国希,高辉远,李鹏民,等. 低温下壳聚糖处理对黄瓜幼苗生理生化特性的影响[J]. 植物生理与分子生物学学报,2004,30(4):441-448.
[23]Xin Z,Browse J. Eskimo1 mutants ofArabidopsisare constitutively freezing-tolerant[J]. Proceedings of the National Academy of Sciences of the United States of America,1998,95(13):7799-7804.
[24]刘春英,陈大印,盖树鹏,等. 高、低温胁迫对牡丹叶片PSⅡ功能和生理特性的影响[J]. 应用生态学报,2012,23(1):133-139.
[25]许 楠,孙广玉. 低温锻炼后桑树幼苗光合作用和抗氧化酶对冷胁迫的响应[J]. 应用生态学报,2009,20(4):761-766.
[26]李春燕,陈思思,徐 雯,等. 苗期低温胁迫对扬麦16叶片抗氧化酶和渗透调节物质的影响[J]. 作物学报,2011,37(12):2293-2298.
[27]何丽斯,汪 仁,孟祥静,等. 茉莉扦插苗对模拟低温的生理响应[J]. 西北植物学报,2010,30(12):2451-2458.
