盐度骤变和渐变对南极大磷虾生存状况和蜕壳的影响
2018-06-29赵国庆邱盛尧黄洪亮李灵智屈泰春
赵国庆, 邱盛尧, 黄洪亮, 李灵智, 屈泰春
(1.烟台大学海洋学院,山东烟台 264005;2.中国水产科学研究院东海水产研究所,农业部东海与远洋渔业资源开发利用重点实验室,上海 200090)
南极大磷虾(EuphausiasuperbDana),隶属于节肢动物门(Arthropoda),甲壳纲(Crustacea),磷虾目(Euphausiidae),磷虾属,其成熟个体体长一般为40~65 mm,个体最大体质量可达2 g,是磷虾种类中数量最多、个体最大的种类[1-5]。南极大磷虾系集群方式生活,主要集中分布于200 m以上水层,且广泛分布于南极大陆架周围各水域,是多年生大型浮游类甲壳动物[6-7]。
最近40年来世界上许多国家对于南极大磷虾的研究十分关注,但是到目前为止南极大磷虾的生物学特性以及生活习性依旧具有一定的不确定性,它们的资源动态也无法被全面掌握[8-9]。由于南大洋特别严酷的自然条件,且考察研究耗资巨大,以及在生产船上受到实验器材和实验空间的限制,关于南极磷虾暂养实验的研究依旧是研究的难点。关于环境因子对于南极磷虾影响的研究多有报道,诸如温度[10]和光照[11]。而盐度是影响水生生物生存的重要环境因子,有节律的降低盐度能够缩短对虾的蜕皮周期、提高蜕皮率、促进生长[12-13]。现已有相关研究提出了盐度对于南极磷虾的行为会产生一定的影响[14],但是由于暂养时间太短,并没有对其死亡情况和蜕壳情况进行总结。为了研究南极大磷虾在不同浓度盐度下的死亡和蜕壳情况,本研究基于上海市科委科研计划项目和中国农业部南极海洋生物资源开发与利用项目,大型加工拖网渔船“龙腾”轮于2017年在南极海域执行生产任务期间,随船对南极大磷虾进行了暂养实验,研究了盐度骤变和渐变对其生存和蜕壳的影响,以期为全面掌握南极大磷虾的生活习性提供参考资料。
1 材料与方法
1.1 实验材料
实验样本采集时间为2017年5月5日,取样地点为南极南舍得兰群岛附近海域(60°11′02″S、58°57′05″W),取样时海水表温为-0.2℃,对活体磷虾进行鉴定,所取磷虾样本均为南极大磷虾,均取自同一网次,采集工具为日本制四片式单船中层磷虾拖网,拖速3~4 kn,拖曳时间20 min,选取活性大、无残缺的活体作为实验对象,体长范围30~45 mm。实验用水全部取自外部自然海水,温度-0.2~0.4 ℃,pH 7.79~8.01,盐度33.8~34.6,暂养实验室的光照强度控制在30~60 lx,暂养期间不做投饵和充氧处理。实验过程中,使用TES-1322A电子照度计对暂养室内的光照进行测定,借助电子盐度酸度计测量盐度和pH,使用电子温度计测量温度。暂养容器为500 mL半透明塑料瓶。稀释用水为船上提供的海水淡化水,配置高浓度海水的盐由笔者通过海水加热蒸发自然生成。
1.2 实验方法
1.2.1 盐度骤变
实验设置14±0.4、18±0.4、22±0.4、26±0.4、30±0.4、34±0.4、38±0.4、42±0.4、46±0.4、50±0.4、54±0.4共11个盐度梯度,以34±0.4为基础盐度,骤变实验将活体磷虾直接转入上述盐度水体中。以500 mL的半透明塑料瓶为实验容器。实验前取55个500 mL半透明塑料瓶,分别装入已经调好的各个盐度梯度的海水400 mL。将取来的虾样装入自然海水中暂养24 h,然后将每个实验容器中各装入2尾鲜活虾,设5个平行样,在静水条件下进行。为了准确记录因为盐度骤变而引起磷虾的突然死亡,更准确的掌握盐度骤变对磷虾的影响,将观察频率分两个层次进行,即第一天每隔3 h观察一次,其后每12 h观察一次,记录磷虾的生活状态以及蜕壳情况。每隔24 h更换相应浓度的新鲜海水,每次换水1/2,并将粪便用滤网滤出。IKEDA等[15]通过实验室研究表明: 南极大磷虾在饥饿状态下可存活200多天,而且在饥饿的状态下南极大磷虾仍然继续进行蜕皮,因此本实验不对磷虾进行饵料投放。BROWN等[16]对南极大磷虾在-1、1和3 ℃的蜕皮时间等参数进行研究发现,1 ℃是最佳的生长温度,所以本实验将温度控制在(1±0.5) ℃。有研究指出,在适宜的温度下,南极大磷虾的蜕壳周期为14 d左右[17],因此,本实验共持续14 d。将试验组与对照组进行对比,蜕壳频率显著增大或者减小的试验组视为盐度的改变对磷虾产生了影响。
1.2.2 盐度渐变
取两个5 L的透明塑料容器,注入盐度为34的海水4 L作为母容器,每个母容器中暂养磷虾60尾。暂养24 h后,取10尾分别放入盐度为34±0.4的5个500 mL半透明塑料瓶中,每个500 mL小瓶中放2尾,共设5个平行样,500 mL小瓶盐度保持不变。母容器盐度升至38±0.4,24 h后,从盐度为38±0.4的母容器中取磷虾10尾,分别放入5个盐度为38±0.4的500 mL小瓶中。母容器盐度再升至42±0.4,适应24 h后,再从母容器中取磷虾10尾分别放入5个盐度为42±0.4的500 mL小瓶中。母容器盐度再升至46±0.4。按此方法,依次将盐度升至54±0.4。为了与骤变组形成对比,观察频率分两个层次进行,即第一天每隔3 h观察一次,其后每12 h观察一次,记录磷虾的生活状态以及蜕壳情况。每隔24 h进行更换相应浓度的新鲜海水,每次换水1/2。盐度渐变速率为4·d-1,即每隔6 h将盐度升高1度。盐度逐渐降低的实验设置与逐渐升高的实验设置方法一致,起始盐度也为34±0.4,逐渐降低至14±0.4。饵料和温度的处理与盐度骤变实验保持一致。实验过程共持续14 d。
1.2.3 盐度渐变后恢复
14 d后,完成盐度逐渐升高和降低实验,将未死亡的磷虾全部放到盐度为34的海水中,培养24 h后,观察记录其生活状态。
1.3 数据统计分析
南极大磷虾的存活率是指各盐度下该组磷虾的存活个数与实验开始时该组磷虾总数的百分比。蜕壳率是指各盐度下磷虾蜕壳的个数与实验开始时该组磷虾总数的百分比。
实验数据用Excel 软件进行处理,采用平均值±标准差(mean ± SD) 表示,数据的统计分析利用SPSS 19.0软件进行单因素方差分析(ANOVA),对南极大磷虾的存活率和蜕壳率进行显著性检验,以P<0.05作为差异显著水平。
2 结果与分析
2.1 盐度骤变对南极大磷虾的影响
2.1.1 存活率
24 h内,盐度骤变对南极大磷虾存活率的影响见表1。由表1可知,盐度骤变显著影响南极大磷虾的存活率(P<0.05)。盐度骤变6 h后,盐度为14时的磷虾个体死亡率达到100%,第12小时死亡率达到100%的骤变盐度组有14±0.4、18±0.4、22±0.4和54±0.4,占总盐度组的36%,而盐度为30~42时,24 h磷虾死亡率为0。而其后在继续暂养的过程中,剩余存活磷虾的死亡率很低,从总体上来看,经单因素分析得出,盐度因素对于磷虾死亡率的影响依然具有显著性(P<0.05)。14 d后,盐度30~42的3个骤变组中,磷虾个体的死亡率依旧为0;而盐度为50±0.4组磷虾全部死亡。说明在盐度骤变时,南极大磷虾可以较好的适应盐度的变化范围为30~42。此外,观察还发现,除盐度50±0.4外,在其它盐度范围内,只要开始适应后,在其后一段时间里,磷虾的死亡率极低。
2.1.2 蜕壳率
24 h内,各盐度组均未出现蜕壳的磷虾个体。在之后的暂养实验中,仅盐度26±0.4、30±0.4、34±0.4、38±0.4和42±0.4组出现了蜕壳现象。盐度为38时蜕壳率最高,占总蜕壳个体的39%。在盐度30~38时磷虾的蜕壳率较高,占磷虾总蜕壳个体的比例为88%(图1)。从盐度骤变实验结果来看,当水体盐度趋于高限时,磷虾的蜕壳速度加快,而当盐度降低低于底限或升高超过高限时,磷虾便不再进行蜕壳。
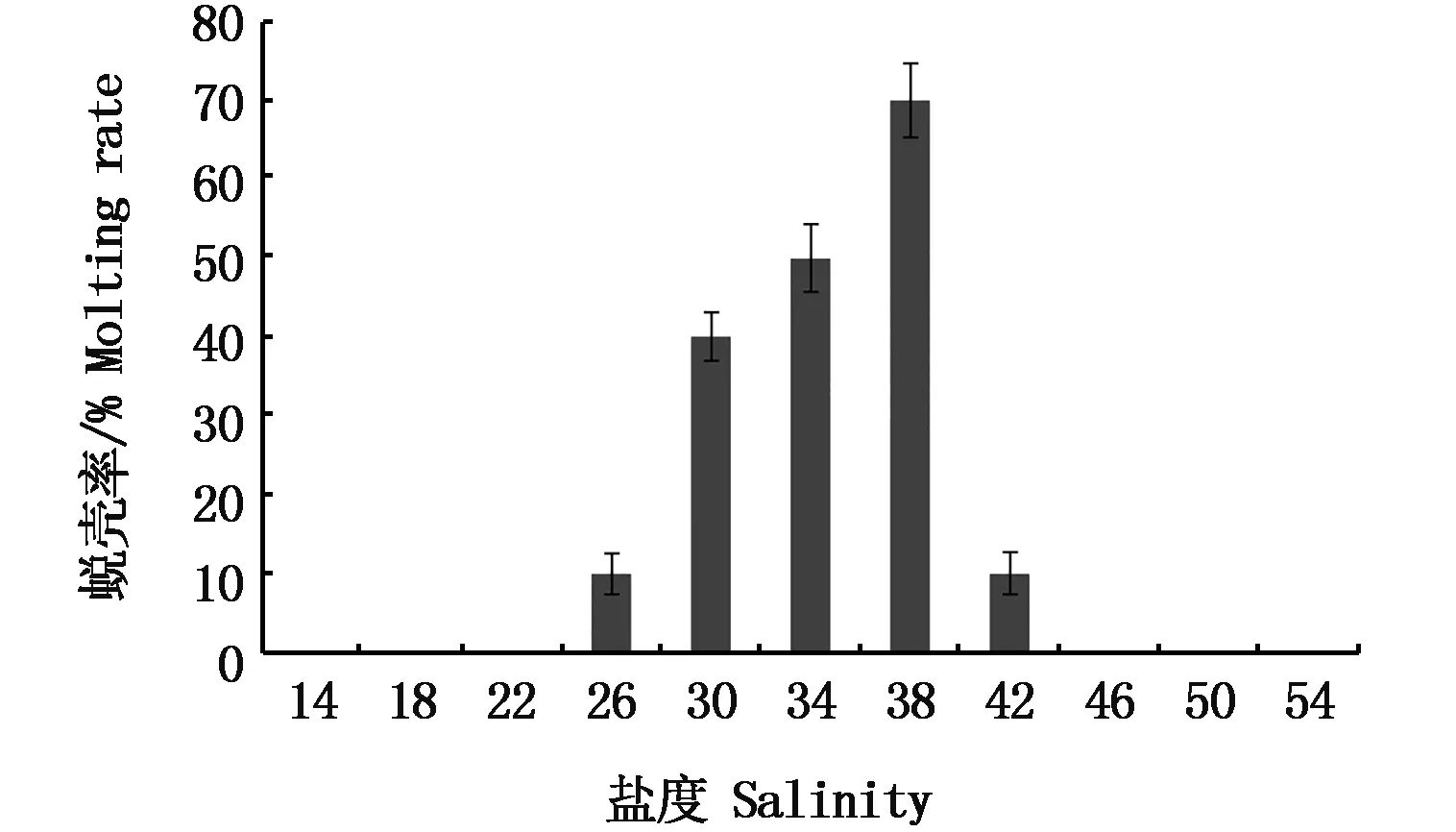
图1 盐度骤变下南极大磷虾的蜕壳率Fig.1 Molting rate of Euphausia superb under sudden change of salinity
2.2 盐度渐变对南极大磷虾的影响
2.2.1 存活率
在母容器暂养过程中,降低组逐渐降到19±0.4、升高组逐渐升到51±0.4时,6 h后进行观察,母容器中的磷虾个体都已全部死亡。24 h内,盐度渐变对南极磷虾存活率的影响见表2。由表2可知,盐度渐变对磷虾的存活率存在显著影响(P<0.05)。24 h内,除暂养已经死掉的14±0.4、18±0.4和54±0.4盐度组外,磷虾的死亡只出现在盐度为22±0.4时,死亡率为20%。盐度升高为50±0.4时,磷虾的活力降低,只能进行腹肢摆动,但是并没有出现死亡情况。当盐度在26~46范围内时,磷虾的死亡率为0,且能够进行正常游动。而其后在磷虾的继续暂养过程中,磷虾的死亡率依然很低,14 d后剩余磷虾的死亡只出现在22±0.4、46±0.4和50±0.4盐度组。而当盐度在26~42范围内渐变时,其死亡率为0。盐度渐变实验表明,磷虾的适宜生长盐度为26~42。
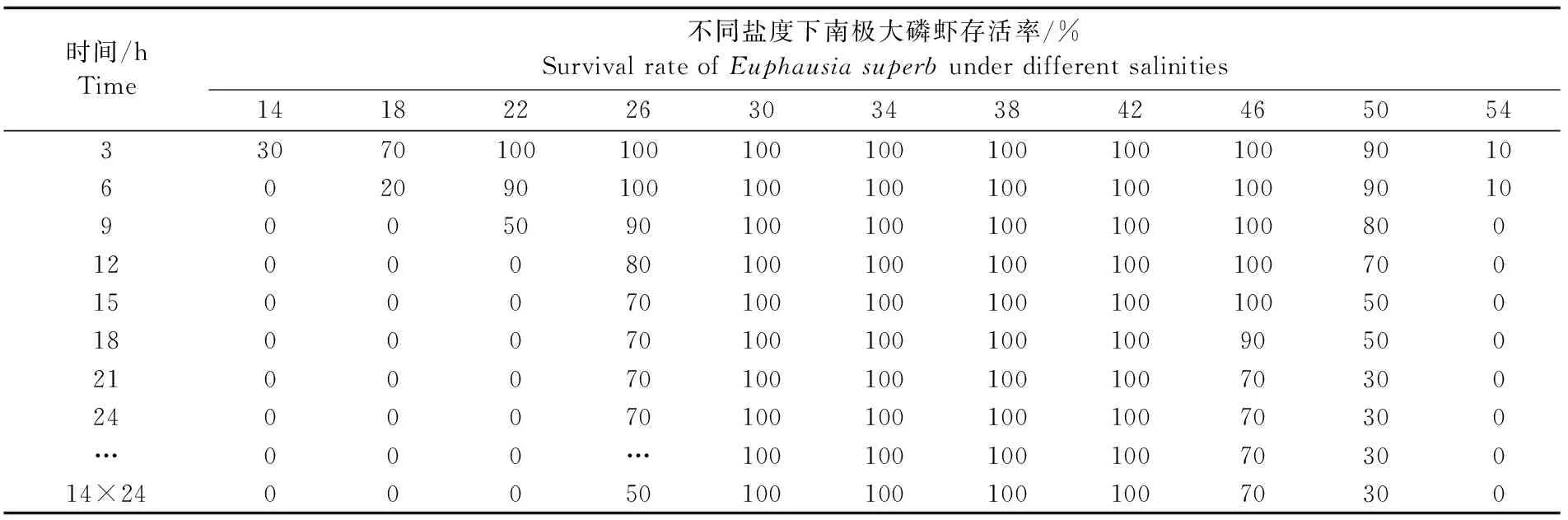
表1 盐度骤变下南极大磷虾的存活率Tab.1 Survival rate of Euphausia superb under sudden change of salinity

表2 盐度渐变下南极大磷虾的存活率Tab.2 Survival rate of Euphausia superb when salinity changes gradually
2.2.2 蜕壳率
在盐度渐变的24 h内,盐度为26±0.4和30±0.4的小组内磷虾都出现了蜕壳现象,蜕壳率分别为10%、20%。在之后的暂养实验中,出现蜕壳的盐度组有26±0.4、30±0.4、34±0.4、38±0.4和42±0.4,蜕壳率分别为10%、20%、50%、70%和20%。盐度为38±0.4时,蜕壳率为70%,达到蜕壳率的最高值。实验观察表明,南极大磷虾的蜕壳主要发生在34±0.4和38±0.4两个盐度组,占总蜕壳个体总数的70%(图2)。
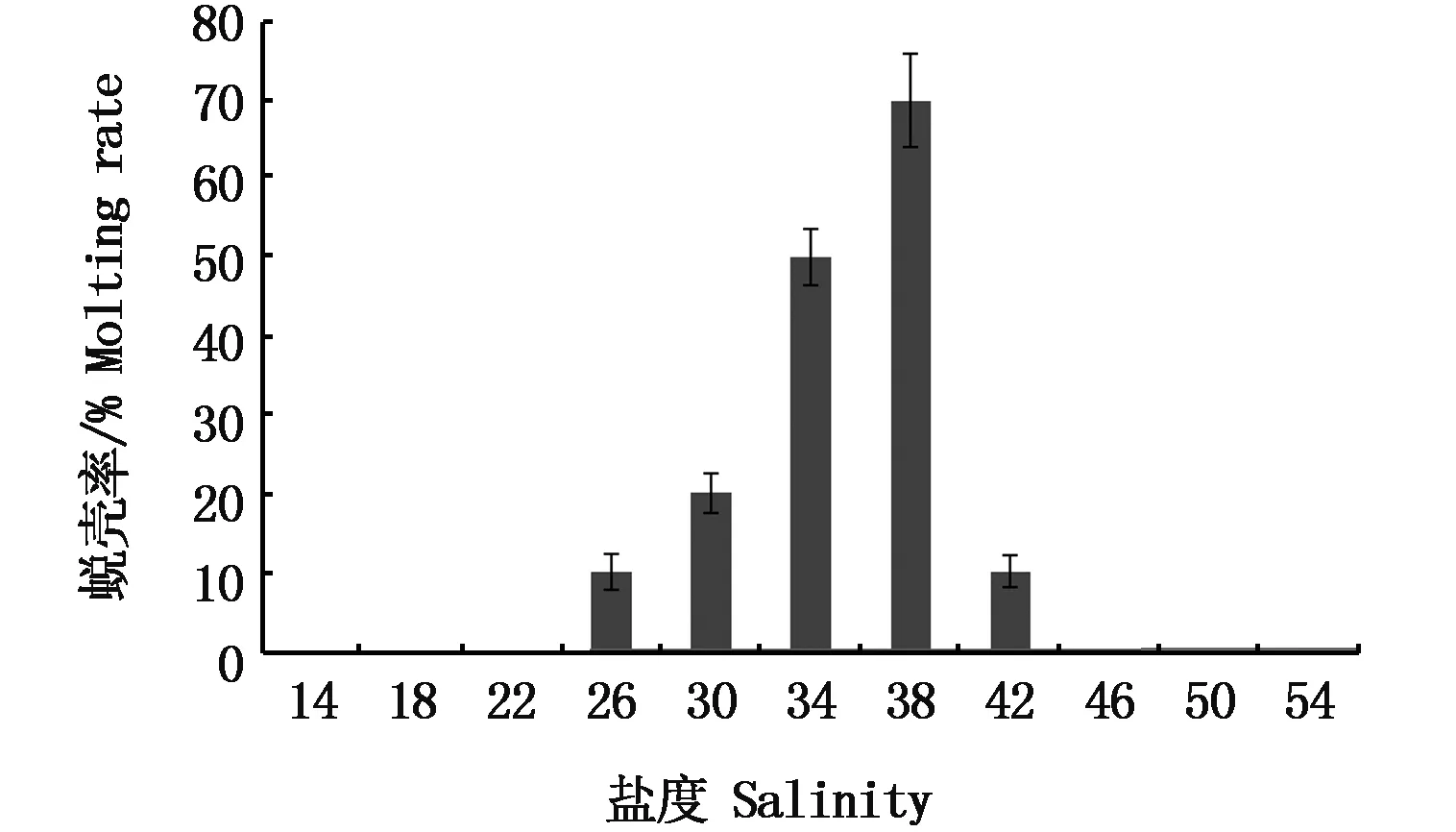
图2 盐度渐变下南极大磷虾的蜕壳率Fig.2 Molting rate of Euphausia superb when salinity changes gradually
2.3 盐度渐变恢复后对南极大磷虾的影响
盐度逐渐升高和逐渐降低的实验后,将所有盐度梯度存活的个体全部转入盐度为34的海水中,培养24 h后,未发现死亡个体,存活率为100%,且亦未出现蜕壳现象。
3 讨论
3.1 南极大磷虾对盐度的适应性
盐度作为海水固有特性,对水生生物的生长发育等生理特性影响较大,不同盐度下的水生生物会表现出不同的生理状态[18]。关于盐度对虾蟹类生长发育的影响的研究很多,丁森等[19]报道,盐度波动幅度过大不利于中国明对虾(Fenneropenaeuschinensis)稚虾的生长;ROMANO等[20]发现,高盐和低盐环境延长了远海梭子蟹(Portunuspelagicus)的蜕壳时间,减少了蜕壳增重,降低了蟹的特定生长率;陈昌生等[21]报道,低盐或高盐对中国龙虾(Panulirusstimpsoni)幼体的生长、发育及蜕皮等有显著影响。朱春华[22]曾对凡纳滨对虾(Penaeusvannamei)进行过研究,结果表明,当盐度低于10或高于26时,为维持自身渗透平衡,凡纳滨对虾需要耗费更多的能量。本研究中,通过盐度骤变和渐变两组实验,通过对盐度进行量化改变,在一定程度上测定了南极大磷虾对盐度的适应性。结果表明,盐度骤变和渐变都显著影响南极大磷虾的存活率(P<0.05)。当盐度在30±0.4到42±0.4范围内时,14 d内南极大磷虾均未出现死亡现象;当盐度为14±0.4、18±0.4和54±0.4时,14 d时南极大磷虾的死亡率均达到100%。由此可以推断,盐度在30~42范围内变化时,南极大磷虾具有较好的适应性。王震等[14]基于海上水箱实验对南极磷虾耐盐性的研究表明,磷虾在水体中正常活动的适宜盐度范围为26~43。本实验的结果与其结论基本一致,但其结果适宜盐度范围要广一些,可能是与本实验时间较长有关。总的来说,南极大磷虾对盐度的变化具有一定的适应能力。
3.2 盐度对南极大磷虾蜕壳的影响
南极大磷虾的重要生物学特点之一是在其一生中不断地进行蜕壳,即使在环境恶劣的情况下也是如此[23],可以说,其生长是通过不断地蜕壳来实现的[24]。南极大磷虾的蜕壳特性很大程度上是为了适应南大洋冬季气温低、饵料不足的条件而存在的,这与其它甲壳类只是为了不断生长、个体不断增大而蜕壳并不类似。IKEDA等[16]、THOMAS等[25]、MCWHINNIE等[26]研究发现,在饵料不足的情况下,磷虾会出现整个身体缩小的现象,有些已经成熟的个体也会通过不断的蜕壳而失去原有的性特征而成为未成体。MAIHARA等[27]、NICOL等[28]研究证明,南极大磷虾的蜕壳周期主要受温度影响。IKEDA等[16]曾经在低温实验室内将南极大磷虾放在过滤海水中培养了211 d,在培养过程中,处于饥饿状态的磷虾伴随着新陈代谢的降低,身体也逐渐缩小,并且在整个实验过程中一直在进行有规律的蜕壳。但是很少有研究者注意并研究盐度对南极大磷虾蜕壳会发生什么样的影响。南极大磷虾在蜕壳过程中,需要与外界环境进行一系列物质循环与能量转换,这不仅需要适宜的盐度来维持渗透压[29],而且需要从外界摄取大量的无机盐如钙、磷、铁、硅等来生成新的外壳[30]。甲壳动物的渗透调节是一种消耗能量的生理过程[22],不同盐度下的渗透调节耗能存在一定的差异[31]。本实验中,在24 h内,盐度骤变组未出现蜕壳个体,而盐度渐变组出现的蜕壳盐度组有两个,分别为26±0.4、30±0.4。在其后的暂养实验中,盐度骤变和盐度渐变中蜕壳率的趋势一致,且出现蜕壳磷虾个体的盐度组也相一致。盐度为14~22和46~54时,均未出现蜕壳个体。当盐度达到38±0.4时,均出现蜕壳率最高值,都为70%,分别占总蜕壳个体的比例为88%、70%。由此可以看出,盐度对磷虾蜕壳率的影响并未因为盐度的骤变和渐变而有所改变,这与盐度对磷虾死亡率的影响并不一致。
在暂养实验过程中发现,在高盐浓度下的南极大磷虾虾壳有变软的现象,这与三疣梭子蟹(Portunustrituberculatus)的暂养实验中高盐抑制了其新壳的硬化现象[32]存在一定的一致性,但是本实验中没有蜕壳的磷虾个体的虾壳也会变软,相关原因还需深入研究。在蟹类里有MDS(molt death syndrome)现象的相关报道,即蟹蜕壳期间旧壳由于无法完全分离而导致的死亡[33]。ROMANO等[34]有过相关报道,远海梭子蟹在盐度10~40下的死亡大多是MDS死亡。但是在本实验中大部分南极大磷虾的死亡似乎并不是因为这个原因,因为在适宜盐度下存活的大部分个体在实验结束前也并未出现蜕壳现象,所以笔者推断在盐度过高和过低的情况下南极大磷虾的死亡可能是因为其渗透调节出现紊乱而导致。
3.3 渐变后恢复对南极大磷虾的影响
盐度渐变后恢复的实验表明,南极大磷虾的成活率为100%。这个结果与其它种类的甲壳类的研究结果相一致,廖永岩等[35]研究表明,中华虎头蟹(Orithyiasinica)从盐度5~55恢复至盐度34时,对存活率影响不显著。关于南极大磷虾的相关研究尚未见报道。在经过长时间一系列盐度变化的培养后,南极大磷虾是否也会相应的增加抗逆境的能力?有关研究还需继续深入探讨。
4 小结与展望
暂养实验对于南极大磷虾环境适应性以及行为学研究具有很重要的指导意义,但是在生产渔船上进行南极大磷虾的暂养实验具有一定的困难,因此目前为止国内关于磷虾现场暂养的研究并不常见。王震等[14]、徐鹏翔等[11]、李莹春等[10]在南极海域捕捞现场分别进行了不同环境因子控制下的南极大磷虾活体暂养实验,研究了南极大磷虾在不同环境因素下的生长、存活状态及耐受性。本实验在方案设计、实验条件、实验材料的获取等方面依然存在一定的缺陷。首先,同一网次活体南极大磷虾的数量受到限制,分组太多导致每一组的磷虾个体数量太少,也无法进行不同成熟级别的分组实验;其次,实验设备较为缺乏,实验场地较为简陋;最后,无法准确地进行个别变量的微调[14]。因此,本实验所得到的结果有可能会略有偏差,各个层面尚有一定改进的空间,但本研究结果也为磷虾环境的适应性研究提供了一定的参考资料。由于南极大磷虾离水后存活时间较短,因此无法将其带至陆地实验室进行暂养实验,海上暂养实验的成功进行,也表明了船上磷虾暂养的可行性。今后在改善船上暂养条件的基础上,可以在各个层面进行南极大磷虾的适应性研究。
参考文献:
[1] 王 荣,孙 松.南极磷虾渔业现状与展望[J]. 海洋科学,1995,19(2):28-32.
WANG R, SUN S. Krill fishery in the southern ocean-its present and future[J].Marine Science,1995,19(2): 28-32.
[2] 黄洪亮,陈雪忠,冯春雷. 南极磷虾资源开发现状分析[J]. 渔业现代化,2007, 34(4): 48-51.
HUANG H L, CHEN X Z, FENG C L. Status ofEuphausuasuperbaresourse exploration[J]. Fishery Modernization, 2007, 34(1): 48-51.
[3] SIEGEL V. Distribution and population dynamics ofEuphausiasuperba: Summary of recent findings[J].Polar Biology,2005,29(1): 1-22.
[4] SIEGEL V,LOEB V,GROGER J. Krill(Euphausiasuperba)density,proportional and absolute recruitment and biomass in the Elephant Island region (Antarctic peninsula) during the period 1977 to 1997[J].Polar Biology,1998,19(6): 393-398.
[5] HAMNER W M, HAMNER P P, STRAND S W,etal. Behavior of Antarctic krill,Euphausiasuperba: chemoreception,feeding,schooling and molting[J].Science,1983(220): 433-435.
[6] 朱国平,冯春雷,吴 强,等.南极磷虾调查CPUE指数变动的影响因素初步分析[J].海洋渔业,2010,32(4):368-373.
ZHU G P, FENG C L, WU Q,etal. Preliminary analysis on factors impacting CPUE index variations in Antarctic krill survey[J]. Marine Fisheries, 2010,32(4):368-373.
[7] 陈雪忠,徐兆礼,黄洪亮. 南极磷虾资源利用现状与中国的开发策略分析[J].中国水产科学,2009,16(3):451-458.
CHEN X Z, XU Z L, HUANG H L. Development strategy on Antartic krill resource utilization in China[J].Journal of Fishery Science of China, 2009,16(3):451-458.
[8] MILLER D G,HAMPTON I. Biology and ecology of the Antarctic krill (EuphausiasuperbaDana) : a review[J]. Biomass Scientific Series,1989(9):1-66.
[9] DALY K L.Overwintering development,growth,and feeding of larvalEuphausiasuperbain the Antarctic marginal ice zone[J]. Limnology and Oceanography,1990,35(7): 1564-1576.
[10] 李莹春,朱国平,孟 涛,等. 人工条件下南极磷虾的温度耐受性实验观察[J].江苏农业科学,2012,40(9): 204-206.
LI Y C, ZHU G P, MENG T,etal. Experimental study on temperature tolerance of Antarctic krill under artificial condition[J]. Journal of Jiangsu Agricultural Sciences, 2012,40(9): 204-206.
[11] 徐鹏翔,李莹春,朱国平,等.光照条件下南极磷虾的行为观察[J].水产学报,2012,36(2): 300-305.
XU P X, LI Y C, ZHU G P,etal. Observation on behaviours of Antarctic krill(Euphausiasuperba) in lighting condition[J]. Journal of Fisheries of China, 2012,36(2): 300-305.
[12] MU Y C,WANG F,DONG S L,etal.The effects of salinity fluctuation in different ranges on intermolt period and growth of juvenileFenneropenaeuschinensis[J].Acta Oceanologica Sinica,2005,27(2):122-126.
[13] BROY W A,LAWRENCE A L,LEUNG-TURGILLO J R.The effect of salinity on growth and survival ofLitopenaeusvannamei,with observations on the interaction of IHHN virus and salinity[J].Aquaculture,1994(122):133-146.
[14] 王 震,朱国平,许柳雄,等.基于现场水箱实验的南极磷虾耐盐性研究[J].大连海洋大学学报,2017,32(2): 211-216.
WANG Z, ZHU G P, XU L X,etal. Saline tolerance of Antarctic krillEuphausiasuperbabased an insitu water tank trail[J]. Journal of Dalian Ocean University, 2017,32(2): 211-216.
[15] IKEDA T, DIXON P. Body shrink age as a possible overw intering mechanism of the Antarcitc krillEuphausiasuperbaDana[J]. Journal of Experimental Marine Biology and Ecology, 1982(62):143-151.
[16] BROWN M,KAWAGUCHI S,CANDY S,etal. Temperature effects on the growth and maturation of Antarctic krill (Euphausiasuperba) [J]. Deep -Sea Research (Ⅱ),2010(57): 672 - 682.
[17] MAIHARA Y, ENDO Y. Laboratory observations on molting and growth of Antarctic krillEuphausiasuperbDana[J]. Memoirs of National Institute of Polar Research, 1986(Special Issue 1):125-127.
[18] FRY F E J. The effect of environmental factors on the physiology of fish[M]//HOAR W S,RANDALL D J. Fish Physiology Vol Ⅵ: Environmental Relations and Behavior. New York: Academic Press, 1971:1-98.
[19] 丁 森,王 芳,郭 彪,等.盐度波动对中国对虾稚虾蜕皮、生长和能量收支的影响[ J].应用生态学报,2008,19(2):419-423.
DING S, WANG F, GUO B,etal. Effects of salinity fluctuation on the molt, growth, and energy budget of juvenileFenneropenaeuschinensi[J]. Chinese Journal of Applied Ecology, 2008,19(2):419-423.
[20] ROMANO N, ZENG C S. The effects of salinity on the survival,growth and haemolymph osmolality of early juvenile blue swimmer crabs,Portunuspelagicus[J]. Aquaculture, 2006,260(14): 151-162.
[21] 陈昌生,陈政强,胡家财,等.中国龙虾叶状幼体的盐度耐受力[J].海洋学报,2003,25(2):24-28.
CHEN C S, CHEN Z Q,HU J C,etal. Study on halotolerance of phyllosoma larva of Chinese spiny lobster (PanulirusstimpsoniHolthuis) [J]. Acta Oceanologica Sinica, 2003,25(2):24-28.
[22] 朱春华. 盐度对南美白对虾生长性能的影响[J]. 水产科技情报,2002,29(4): 166-168.
ZHU C H. Effects of salinity on growth performance ofPenaeusvannamei[J]. Fisheries Science & Technology Information, 2002,29(4): 166-168.
[23] 孙 松,王 荣. 南极磷虾的生长与复眼晶椎数目关系的研究[J],南极研究,1995,7(4):2-6.
SUN S, WANG R. Study on the relationship between the crystalline cone nember and the growth of the Antarctic krill[J]. Antarctic Research(Chinese edition), 1995,7(4):2-6.
[24] 孙 松,王 荣.南极磷虾年龄鉴定研究简述,南极研究,1995,7(2):59-62.
SUN S, WANG R. Aging the Antarctic krill[J]. Antarctic Research(Chinese edition), 1995,7(2):59-62.
[25] THOMAS P G, IKEDA T. Sexual regression, shrinkage, re-maturation and growth of spent femaleEuphausiasuperbain the laboratry[J].Marine Biology,1987(95):357-363.
[26] MCWHINNIE M A, DENYS C J. The high importance of lowly krill[J].Natural History,1980(89): 66- 73.
[27] MAIHARA Y, ENDO Y. Laboratory observations on molting and growth of Antarctic krillEuphausiasuperbDana[J]. Memoirs of National Institute of Polar Research,1986(Special Issue 1):125- 127.
[28] NICOL S, STOLP M. A refinement of the moult staging technique for Antarctic krill(Euphausiasuperba) [J] .Marine Biology,1990(104):169-173.
[29] 李二超.盐度对凡纳滨对虾的生理影响及其营养调节[D].上海: 华东师范大学,2008.
LI E C. Physiological effects of ambient salinity onLitopenaeusvannameiand nutrient modulation[D].Shanghai: East China Normal University,2008.
[30] 杨文杰,许柳雄.环境因素对南极磷虾资源分布影响的研究进展[J].大连海洋大学学报,2014,29(3): 316-322.
YANG W J, XU L X. A review: research progress on environmental factors affecting resource distribution of Antarctic krill[J]. Journal of Dalian Fisheries University,2014,29(3): 316-322.
[31] 吕慧明, 徐善良. 虾蟹能量收支的特点及其影响因素[J]. 水产科学, 2009, 28(10): 604-608.
LV H M, XU S L. Characteristics of energy budget and affecting factors in shrimps and crabs[J]. Fisheries Science, 2009, 28(10): 604-608.
[32] 路允良,王 芳,赵卓英,等.盐度对三疣梭子蟹生长、蜕壳及能量利用的影响[J]. 中国水产科学,2012, 19(2): 237-245.
LU Y L, WANG F, ZHAO Z Y,etal. Effects of salinity on growth, molt and energy utilization of juvenile swimming crabPortunustrituberculatus[J]. Journal of Fishery Science of China,2012, 19(2): 237-245.
[33] YE L, JIANG S G, ZHU X M,etal. Effects of salinity on growth and energy budget of juvenilePenaeusmonodon[J]. Aquacluture,2009(290): 140-144.
[34] ROMANO N, ZENG C S. The effects of salinity on the survival,growth and haemolymph osmolality of early juvenile blue swimmer crabs,Portunuspelagicus[J]. Aquaculture, 2006,260(14): 151-162.
[35] 廖永岩,吴 蕾,蔡 凯,等.盐度和温度对中华虎头蟹(Orithyiasinica)存活和摄饵的影响[J].生态学报,2007,27(2):627-639.
LIAO Y Y, WU L, CAI K,etal. The effect of salinity and temperature on survivorship and food intake of tiger crab,Orithyiasinica[J].Acta Hydrobiologica Sinica, 2007,27(2):627-639.
