基于环境因子的日本鲭太平洋群系的亲体—补充量模型比较分析
2018-06-29林琴琴陈新军戴黎斌
林琴琴, 陈新军,2,3,4,5, 戴黎斌
(1.上海海洋大学海洋科学学院,上海 201306; 2. 农业部大洋渔业开发重点实验室,上海 201306;3.国家远洋渔业工程技术研究中心,上海 201306;4.大洋渔业资源可持续开发教育部重点实验室,上海 201306; 5. 农业部大洋渔业资源环境科学观测实验站,上海 201306)
日本鲭(Scomberjaponicus)属大洋暖水性中上层鱼类,广泛分布于西北太平洋沿岸水域,主要为中国、日本、韩国等灯光围网船所利用。在日本,日本鲭是最主要的鱼类资源之一,分为太平洋群系和对马群系两个种群[1]。太平洋群系日本鲭的总生物量在1978年达到高峰,自1990年以来,日本鲭补充量呈不断下降的趋势,2000年之后,渔获量一直处于较低水平[2],资源状况堪忧,产生这种现象的主要原因是在补充量持续较低的年份进行过度捕捞[3],因此,对太平洋群系日本鲭进行合理的种群管理十分必要。
亲体-补充量关系(stock and recruitment relationship, SRR)是渔业资源评估和管理的基础[4],也是制定渔业资源管理对策的主要依据之一。渔业生物的亲体量(spawning stock biomass,S)对种群的补充量(recruitment,R)有着决定性的影响,然而,由于补充量短时期内的剧烈波动、亲体量的测量误差等原因常使得模型的参数估计存在着很大的不确定性[5]。SAKURAMOTO[6]认为,将环境因子引入SSR模型对于更好的掌握鱼类种群动态是十分必要的。李纲等[7]分析了SST对东、黄海日本鲭资源和持续产量的影响,发现SST是影响日本鲭生长、繁殖和补充等最重要的因素之一。陈新军等[8]利用卫星遥感图像及灯光围网渔业生产统计数据, 探讨日本鲭渔场分布与海表面温度、叶绿素a 浓度和海表面高度之间的关系,表明将3个环境因子进行综合分析,预报潜在渔场的位置更为可靠。
Ricker模型对环境的响应比较显著[9],且已被广泛应用于日本沙丁鱼(Sardinopssagax)、鳀(Engraulisjaponicas)等鱼类种群动态研究中。因此,本文根据1982-2014年日本鲭的生产统计数据,利用Ricker模型探究SST、PDO对日本鲭亲体-补充量的影响,获得最优模型,以把握环境变化对日本鲭资源变动产生的影响,为渔业生产和管理提供参考。
1 材料与方法
1.1 数据来源
本研究所采用的日本鲭亲体量和补充量数据来源于日本中央水产研究所(表1),时间跨度为1982-2014年。
根据YATSU等[3]的研究,采用水表面温度(sea surface temperature,SST)和太平洋十年涛动(Pacific decadal oscillation,PDO)作为表征影响日本鲭亲体-补充量关系的环境因子。SST数据来源于网站http://iridl.ldeo.columbia.edu/SOURCES/.IGOSS/.data_products.html。PDO数据来源于日本气象厅http://www.jma.go.jp/jma/index.html 。海域为129°~160°E、28°~45°N,空间分辨率为1°×1°,时间分辨率为月。
1.2 研究方法
1.2.1 交相关分析
运用R软件,对补充量R与SST、PDO做交相关分析(cross correlation),从而得出SST、PDO变化对补充量R的提前或滞后性影响。交相关分析使用的是时间序列分析,对比两个指数的时间序列,从而得出一个时间序列与另一个时间序列的相关性大小,通常可将分析结果分为时间延迟、时间提前以及变化趋势基本相同或者相反的情况[10]。
1.2.2 SRR模型
Ricker的SRR模型表达式为[11]:
R=αS×e-βS
式中,α、β为模型中的待定参数,R为补充量,S为亲体量。参数α是不依赖群体的指数即与非密度依赖死亡有关的参数,而参数β是依赖于亲体死亡的指数即与密度依赖死亡有关的参数。
为使用GLM函数分析环境对补充量R的影响,对Ricker模型进行优化,将其转换成对数形式的亲体-补充量模型:
ln(R)-ln(S)=ln(α)-βS+θV
式中,V表示环境因子,θ为环境因子系数。将SST、PDO分别加入模型中进行拟合,之后同时加入这两种环境因子构建模型进行分析。
在统计学上,AIC(Akaike information criterion)作为衡量模型拟合优良性和模型选择的标准已经得到广泛的应用[12]。本文根据AIC值,比较Ricker模型与加入环境变量的Ricker模型的拟合效果。其中,AIC值由R软件计算得出,AIC值最小的模型为最优模型。
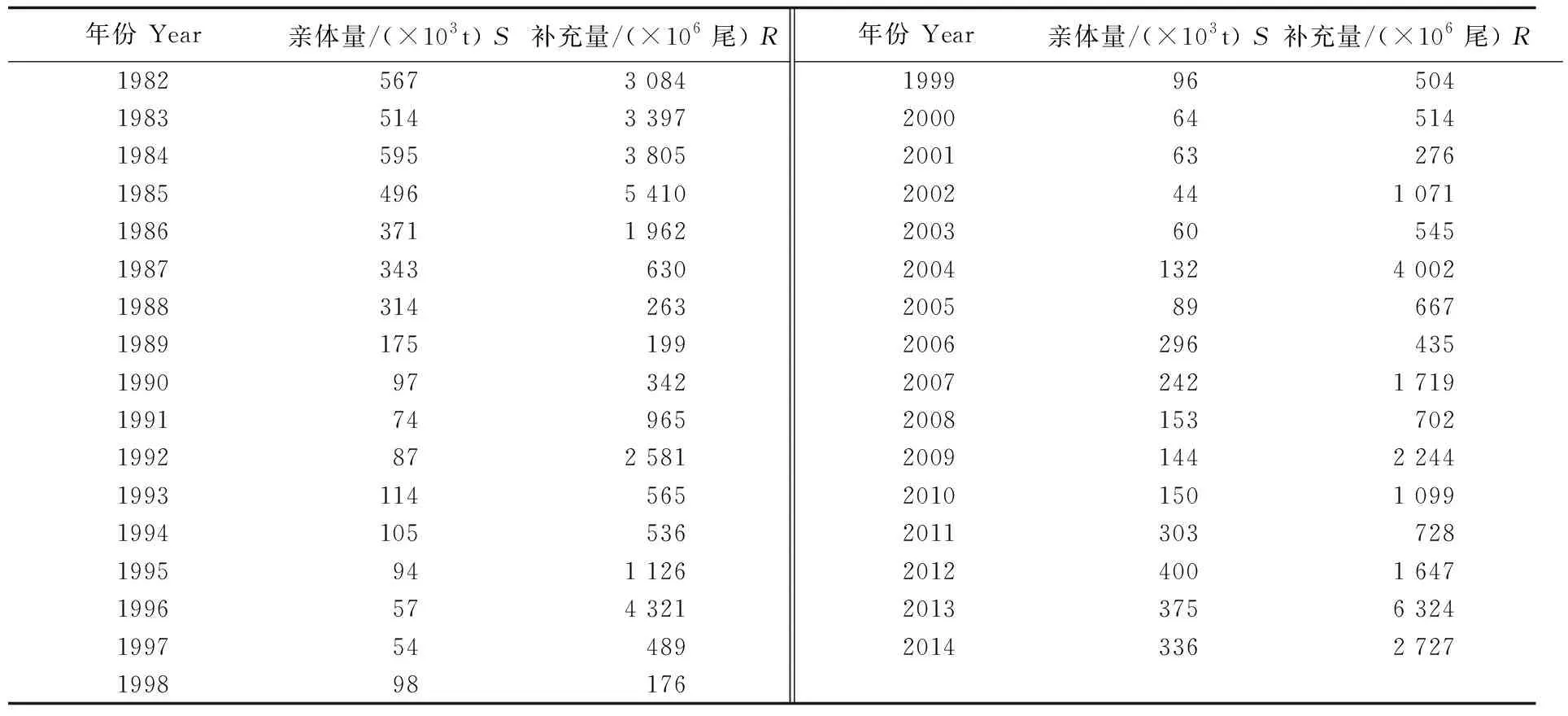
表1 日本鲭太平洋群系的亲体量与补充量数据Tab.1 Spawning stock biomass and recruitment data of Scomber japonicus
2 结果与分析
2.1 交相关分析结果
根据表1所示的1982-2014年日本鲭亲体量(×103t)与补充量(×106尾)资料,用R软件进行交相关分析,图1为输出结果。图1中,横轴数值表示延迟或提前的时间(以年为单位),正值表示延迟,负值表示提前。纵轴数值表示两者相关性大小,通常绝对值数值越高表示相关性越高,反之,则相关性越低,正、负值分别代表两者呈正相关和负相关。由图1可看出,补充量R与PDO的相关系数最大的为提前一年的PDO,为正相关,表明提前1年的PDO是描述补充量变化的最适模型;补充量R与SST的相关系数最大的为滞后5年的SST,为正相关,表明滞后5年的SST是描述补充量变化的最适模型。
2.2 基于不同环境因子的Ricker模型
从图2可以看出,和大多数鱼类一样,日本鲭亲体量与补充量的数据分布广泛,个别数据非常零散,大致符合Ricker模型的分布趋势。根据时间序列分析结果,将PDO数据提前1年,将SST数据延迟5年,分析1987-2014年的亲体-补充量数据,用GLM函数构建加入不同环境因子的Ricker模型。对比4种Ricker模型(表2),单纯用Ricker模型进行拟合的AIC值为81.979;加入PDO的Ricker模型的AIC值为77.566,P值为0.018;加入SST的Ricker模型的AIC值为74.941,P值为0.005;加入PDO、SST两种环境因子的Ricker模型的AIC值为71.743,较之单纯用Ricker模型、加入环境因子的Ricker模型的拟合效果更好。其中,SST对补充量变化的影响比PDO更显著,加入SST模型的拟合效果要优于加入PDO的拟合效果。加入PDO、SST两种环境因子的Ricker模型的AIC值最小,拟合效果最好,模型公式为: ln(R)-ln(S)=-11.936+0.005S-0.394VPDO+ 0.841VSST,这表明环境因子对日本鲭亲体-补充量关系有显著影响。

图2 日本鲭亲体量与补充量数据散点图Fig.2 Scatter plot of spawning stock biomass and recruitment data of Scomber japonicus

图1 PDO、SST与补充量R的交相关分析结果Fig.1 Results of cross correlation analysis between PDO, SST and recruitment

模型Model系数Coefficient估计值Estimate标准误差Std.errort值t valueP值P valueR~S截距Intercept2.518-0.3287.6723.84×10-8斜率Slope-0.0040.002-2.3710.025AIC81.979R~S+PDO截距Intercept2.6470.3038.7434.49×10-9S系数Coefficients of S-0.0040.002-2.8170.009PDO系数Coefficients of PDO-0.4970.196-2.5370.018AIC77.566R~S+SST截距Intercept-14.4615.509-2.6250.014 6S系数Coefficients of S-0.0050.001-3.3140.003SST系数Coefficients of SST0.9810.3183.0860.005AIC74.941R~S+PDO+SST截距Intercept-11.9365.250-2.2730.032S系数Coefficients of S-0.0050.001-3.6530.001PDO系数Coefficients of PDO-0.3940.178-2.2120.037SST系数Coefficients of SST0.8410.3022.7810.010AIC71.743
3 讨论
3.1 PDO 与 SST 的滞后性影响
PDO是一种10年周期尺度变化的太平洋气候变化现象,变换周期通常为20~30年。PDO的特征为在 “暖相位”期间西太平洋偏冷,而在“冷相位”期间西太平洋偏暖。黑潮是沿北太平洋西部边缘向北流动的一支强西边界海流,黑潮的发生与PDO有密切关系,而黑潮又是引起SST出现年际变化的重要因素,SST的空间分布及其增减与黑潮势力的强弱有密切关系[12]。COLLIE等[13]将PDO、黑潮等气候现象定义为一种在海洋条件下的低频率、高振幅的变化,主要在生物量的变动上表现出来,可通过几个营养级进行传播。国外学者的许多研究也证实了SST、厄尔尼诺现象、南方涛动等因素对于日本鲭资源丰度的影响[14-15]。通常情况下我们认为,在气候发生变化的一定时间以后,才会对日本鲭产卵场及索饵场的SST产生影响,进而影响到渔业资源丰度。本研究中,根据交相关分析结果,提前1年的PDO和滞后5年的SST会对日本鲭补充量变化产生显著性影响。日本鲭的主要产卵时间为每年的3-6月,6-10月仔稚鱼随水流洄游至索饵场,之后在此越冬。性成熟年龄为2龄,少数鱼在1龄也可以性成熟,这些未成熟鱼是日本鲭的重要补充群体。日本鲭产卵量和仔幼鱼的存活率受温跃层深度、氧浓度、营养成分等条件的影响,而PDO现象提前发生1年会给仔稚鱼带来适宜的外界环境条件。日本鲭最大寿命为18龄[16],生殖群体一般为2~7龄,可重复产卵。日本鲭的个体绝对生殖力随年龄的增长而增加,在高龄鱼中雌鱼所占比例较高,5~7龄的产卵概率可达到90%以上[17]。滞后5年的SST是利于日本鲭生长的适宜温度,生长迅速,而此时日本鲭的亲体量较高,从而影响日本鲭的资源变动。OH等[18]的研究表明,在产卵季节,推迟3~5年产卵场的SST可以很好的解释日本海的狭鳕渔获量的波动,这和本文得出的滞后5年的SST会对亲体-补充量关系变化产生显著性影响是一致的。PDO和SST的这种提前和滞后效应对研究日本鲭的种群动力机制及预测资源丰度的变化趋势有很大帮助。
3.2 PDO 、SST 对亲体-补充量关系的影响
本文的主要目的是研究环境因子是否对亲体-补充量关系有显著影响,根据这一观点,本研究选取了PDO、SST两个影响因子,从而构建了4种Ricker模型,通过对4种Ricker模型拟合优度的比较,加入PDO、SST两种环境因子的Ricker模型AIC值最小,拟合效果最好,这与前文提到的综合多种环境因子对渔场进行预测所获得的结果更可靠是一致的。
从上述分析结果可知,PDO对日本鲭补充量变化有显著性影响。日本鲭以小型深海生物为食,主要是浮游生物和无脊椎动物。日本鲭游泳能力强,每年进行季节性洄游,春夏时多栖息于水温和盐度较高的海域进行产卵索饵。PDO是黑潮、厄尔尼诺现象等几种不同海洋气候变化过程的综合结果,而不是某种单一的物理模态现象[19]。不同性质的水团交汇,使得浮游生物量增加,为鱼群提供了丰富的饵料和其他营养物质。而黑潮水体在海洋锋附近海域的剧烈混合,形成了鱼群索饵洄游的良好通道。李曰嵩等[20]等采用物理环境因子,建立了日本鲭早期生长史模型,发现在黑潮锋面靠近暖水一侧附近会有大量幼鱼分布,表明外界环境会对仔幼鱼的输运和补充产生影响。
根据表2的拟合效果,SST对补充量变化的影响比PDO更显著,表明日本鲭的补充量很大程度上受SST的直接影响,和PDO相比,SST提供了更多有关种群动态变化的重要信息。SST会对日本鲭的产卵成功率和早期存活率造成很大的影响,而仔稚鱼是重要的补充群体。黑潮-亲潮过渡区域是日本鲭最主要的产卵场,亲潮水流的大量涌入改善了此区域的营养条件,促进了幼鱼的生长[20-22]。NISHIDA[23]通过分析Ricker模型中亲体与补充量的残差,发现海表面温度越高的区域补充量越高。HWANG[24]也指出,在水温相对较高的6-8月,0岁鱼的丰度较大。从亲体量与补充量的拟合曲线图(图3)可看出,和观测数据相比,加入环境因子的拟合曲线更加符合Ricker模型的趋势。加入SST、PDO两种环境因子的拟合曲线在后半段出现上升现象,这可能是由于个别数据较分散造成的,无法全面反映补充量的变化趋势,关于综合多种环境因子对亲体-补充量关系影响的研究还应进一步深入。
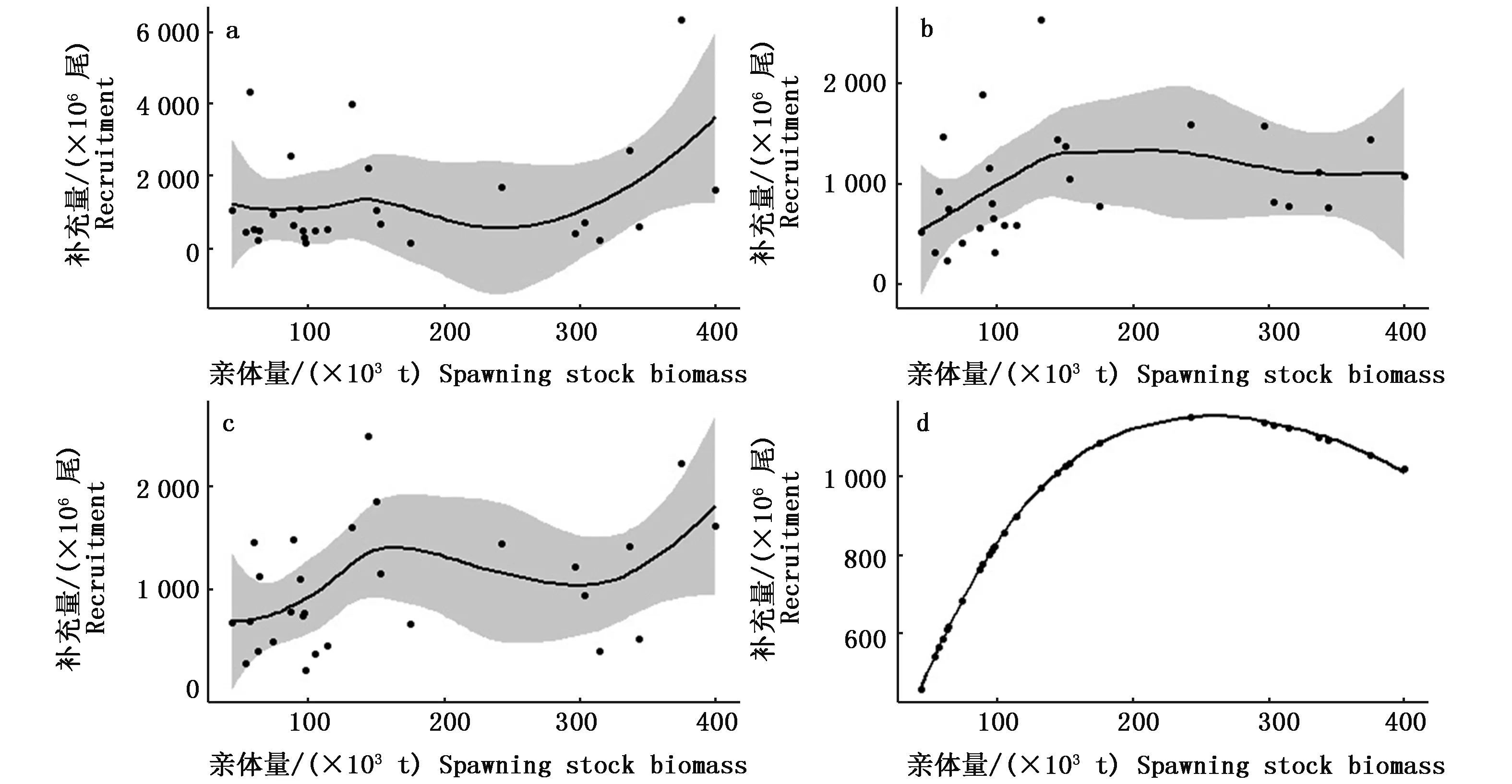
图3 亲体量与补充量的拟合曲线图(a表示观测数据的拟合曲线,b表示加入SST的Ricker模型的拟合曲线,c表示加入SST、PDO的Ricker模型的拟合曲线,d表示Ricker模型的拟合曲线, 阴影部分表示95%置信区间)Fig.3 Fitting curve of spawning stock biomass and recruitment(a represents the fitting curve of observed statistics, b represents the fitting curve of Ricker model with SST, c represents the fitting curve of Ricker model with SST and PDO, d represents the fitting curve of Ricker model. The shaded area represents the 95% confidence limit)
近年来,日本鲭年龄结构趋向低龄化,群体结构趋向简单化,个体小型化现象明显,种群已处于生长型捕捞过度状态[25]。了解气候变化在年际和年代际时间尺度上对鱼类补充量的影响是了解鱼类种群动态和海洋生态系统的基础[26]。因此,为把握环境变化对日本鲭资源变动产生的影响,应进一步加强对日本鲭行为学的研究,综合其他环境因子加以分析,为渔业生产提供更为丰富的科学依据。
参考文献:
[1] WATANABE C, YATSU A. Long-term changes in maturity at age of chub mackerel (Scomberjaponicus) in relation to population declines in the waters off northeastern Japan[J]. Fisheries Research, 2006, 78(2):323-332.
[2] YUKAMI R, OHSHIMO S, YODA M,etal. Estimation of the spawning grounds of chub mackerelScomberjaponicus, and spotted mackerelScomberaustralasicus, in the East China Sea based on catch statistics and biometric data[J]. Fisheries Science, 2009, 75(1):167-174.
[3] YATSU A, WATANABE T, ISHIDA M,etal. Environmental effects on recruitment and productivity of Japanese sardineSardinopsmelanostictus, and chub mackerelScomberjaponicus, with recommendations for management[J]. Fisheries Oceanography, 2005, 14(4):263-278.
[4] RICKER W E. Computation and interpretation of biological statistics of fish populations [J]. Bulletin of Fisheries Research Board of Canada, 1975, 191(18): 1-382.
[5] HILBORN R, WALTERS C J. Quantitative fisheries stock assessment: Choice, dynamics and uncertainty [M]. New York : Chapman & Hall, 1992.
[6] SAKURAMOTO K. Does the Ricker or Beverton and Holt type of stock-recruitment relationship truly exist?[J]. Fisheries Science, 2005, 71(3):577-592.
[7] 李 纲, 郑晓琼, 朱国平,等. 基于水温因子的东、黄海鲐鱼剩余产量模型建立[J]. 上海海洋大学学报, 2011, 20(1):108-113.
LI G, ZHENG X Q, ZHU G P,etal. Establishment of surplus production model for chub mackerel (Scomberjaponicus)in the East China Sea and Yellow Sea based on the sea surface temperature[J]. Journal of Shanghai Ocean University, 2011, 20(1):108-113.
[8] 李 纲, 陈新军. 夏季东海渔场鲐鱼产量与海洋环境因子的关系[J]. 海洋学研究, 2009, 27(1):3-10.
LI G, CHEN X J. Study on the relationship between catch of mackerel and environmental factors in the East China Sea in summer[J]. Journal of Marine Sciences, 2009, 27(1):3-10.
[9] ZHAO X, HAMRE J, LI F,etal. Recruitment, sustainable yield and possible ecological consequence of the sharp decline of the anchovy (Engraulisjaponicus) stock in the Yellow Sea in the 1990s[J]. Fisheries Oceanography, 2003 ,12(4): 495-501.
[10] DAVIS R A JR. Oceanography: An introduction to the marine environment[M]. Wiley, 1970:783-789.
[11] RICKER W E. Stock and Recruitment[J]. Journal of the Fisheries Research Board of Canada, 1954, 11(5) :559-623.
[12] 郑晓琼, 李 纲, 陈新军. 基于环境因子的东、黄海鲐鱼剩余产量模型及应用[J]. 海洋湖沼通报, 2010(3):41-48.
ZHENG X Q, LI G, CHEN X J. Application of environmentally dependent surplus production model forScomberjaponicusin the East China Sea and Yellow Sea[J]. Transactions of Oceanology and Limnology, 2010(3):41-48.
[13] COLLIE J, RICHARDSON K, STEELE J. Regime shifts: Can ecological theory illuminate the mechanisms?[J]. Progress in Oceanography, 2015, 60(2): 281-302.
[14] HIYAMA Y, YODA M, OHSHIMO S. Stock size fluctuations in chub mackerel (Scomberjaponicus) in the East China Sea and the Japan East Sea[J]. Fisheries Oceanography, 2002, 11(6):347-353.
[15] SUN C H, CHIANG F S, SOAC E T,etal. The effects of El Nio on the mackerel purse-seine fishery harvests in Taiwan: An analysis integrating the barometric readings and sea surface temperature[J]. Ecological Economics, 2006, 56(2):268-279.
[16] 川端 淳, 渡邊千夏子, 由上龍嗣. マサバ太平洋系群·対馬暖流系群の資源の長期変動と近年の動態[J]. 水産海洋研究, 2012(76):225-227.
KAWABATA J, WATANABE C, YUKO R. Long-term fluctuation of resources of Massaba Pacific group · Tsushima warm current group and recent dynamics [J]. Marine Research on Oceanography, 2012(76): 225-227.
[17] 刘 松, 顾晨曦, 严 正. 鲐鱼个体生殖力研究[J]. 海洋科学, 1988, 12(5):43-48.
LIU S, GU C X, YAN Z. A study on the individual fecundity of mackerel,Pneumatophorusjaponicus[J]houttuyn, Marine Sciences, 1988, 12(5):43-48.
[18] OH T, SAKURAMOTO K, HASEGAWA S. On the relationship between water temperature and catch fluctuations of walleye pollack Theragra chalcogramma in the northern waters of the Japan Sea[J]. Nippon Suisan Gakkaishi, 2002(68): 866-873.
[19] PHELPS CARLY. Effects ofPDOregime on summer melt of glaciers in the Cascades[D]. University of Delaware, 2012.
[20] 李曰嵩, 潘灵芝, 陈新军,等. 东海鲐鱼(Scomberjaponicus)仔幼鱼游泳行为对输运和补充量的影响[J]. 海洋与湖沼, 2013, 44(2):318-325.
LI Y S, PAN L Z, CHEN X J,etal. Impact of swimming behavior on transport and recruitment of larvae and juvenile of chub mackerel (Scomberjaponicus) in East China Sea[J]. Oceanology Et Limnologia Sinica, 2013, 44(2):318-325.
[21] ODATE S. Study on the larvae of the fishes in the north-eastern sea area along the Pacific coast of Japan[J]. Deep Sea Research & Oceanographic Abstracts, 1961(19):98-108.
[22] WATANABE Y, HIYAMA Y, WATANABE C,etal. Inter-decadal fluctuations in length-at-age of Hokkaido-Sakhalin herring and Japanese sardine in the Sea of Japan[J]. Pices Scientific Report, 2002(20):63-67.
[23] NISHIDA H. Long-term fluctuations in abundance of the chub mackerel and jack mackerel immigrants in the western Japan Sea[J]. Bulletin of the Japanese Society of Fisheries Oceanography, 1997(61):316-318.
[24] HWANG S D. Population ecology of pacific mackerelScomberjaponicus, off Korea[D]. Taejon: Chungnam National University (in Korean), 1999.
[25] 郑 波. 鲐鱼年龄和生长及其资源丰度与环境因子关系的研究[D]. 上海:上海海洋大学, 2007.
ZHENG B. Age and growth of mackerel(Scomberjaponicus) and the relationship between its abundance and environmental variables[D]. Shanghai:Shanghai Ocean University, 2007.
[26] BRANDER K. Impacts of climate change on fisheries[J]. Journal of Marine Systems, 2010(79):389-402.
