米老排优树单亲子代测定及优良家系选择
2018-06-27洪永辉林能庆黄钦忠
洪永辉,林能庆,黄钦忠
(1.福建省龙岩市林业种苗站,福建 龙岩 364000;2.福建省上杭白砂国有林场,福建 龙岩 364205)
米老排(Mytilarialaosensis)属金缕梅科常绿乔木,天然分布于我国云南南部、广西西南部和广东西部,是我国南亚热带集材用、生态保护和观赏于一体的重要生态经济树种,也是马尾松中幼林近自然改培针阔混交林首选树种之一,其地位在林业中越来越重要。自20世纪60年代以来,福建省开展米老排引种驯化研究工作,至今在全省推广种植面积达700多hm2,但米老排的种植规模和推广速度均偏小,其主要原因是缺少良种和优良繁殖材料。开展优树选择和子代测定可为营建种子园和开展无性繁殖提供优良材料,也是林木遗传改良有效手段和良种选育的基础。
1 材料与方法
试验地位于福建省上杭白砂国有林场丰面桥工区,116°21′18″ E,25°08′ N,地处武夷山脉南段东坡玳帽山延伸的低山丘陵地段带,土壤均为红壤土类,Ⅳ类地,海拔高200 m,植被为矮芒萁。气候属中亚热带季风气候,夏长不酷热,冬短不严寒,干湿季节明显,雨量相对集中。年平均气温19.9 ℃,年积温7 288 ℃,7月平均气温27.9 ℃,1月平均气温10 ℃,极端最高气温39.7 ℃,最低气温-5 ℃;年降水天数160 d,年降雨量1 605.4 mm,无霜期301 d,年均相对湿度77%。
试验地于2014年冬劈杂灌、开平台,在平台上定点挖穴,穴规格60 cm×40 cm×40 cm,明穴,回表土,株行距3 m×3 m。面积2 hm2。2013年冬对米老排母树林12个优树分家系采种[1],2014年分家系大田育苗[2]。造林按随机区组设计,重复6次,4株小区,试验设计2个对照。其中:CK1为优树混合种子,CK2为母树林混合种子。同时为核实各家系不同海拔造林适应性和抗逆性,选择海拔600 m,Ⅱ类地九岗工区山顶分家系造林。2017年9月初调查树高、胸(地)径、冠幅等性状指标,采用Excel和SAS统计软件进行数理统计和遗传分析[3]。
2 结果与分析
2.1 米老排造林关键技术研究及家系成活率调查
米老排为浅根性阔叶树种,木材为散孔材,其树叶面积大且浓绿,幼苗主要特征是茎秆髓心为空洞,且空洞部分占断面积50%以上,而植物营养和水分传输主要依靠木质部的导管,同其他散孔材阔叶树相比,米老排木质部因髓心空洞造成营养和水分传输更薄,树叶面积大,其蒸腾作用更强,苗木移植时若植物根系吸水赶不上叶片蒸腾必将造成苗木萎蔫,在春季树木未萌动前完成米老排苗木移植是保持苗木水分平衡提高造林成活率关键。丰面桥调查表明,米老排造林成活率仅为71.8%,各家系成活率为53.%~95%,成活率按由高至低排序为2>1>6>9>3、4、5、10>7>8>12>CK1>CK2。其中成活率大于85%的是2、1、6、9,而3、4、5、10成活率在75%以上。分析可知,3月中旬立春和雨水已过,造林时间太迟,苗木平均高在1.07m以上且顶芽已经萌动[2],虽将苗木树叶摘除只保留顶芽。但因祼根苗造林后遭遇连续干旱、高温是成活率降低主要因素。九岗造林成活率为71.3%,该试验点位于海拔600 m山顶风口,常遭受7级以上大风或台风影响,7、8、9家系出现树木折断后从基部重新萌芽。但2号家系造林成活率达100%,8、9号家系成活在85%以上,其生长表现良好,综合说明上述家系对环境有较强的适应性,其抗逆性和适应性较强。
2.2 米老排各试验点生长性状表现
米老排幼龄期均表现为速生(表1),其12个优树单亲子代平均树高、胸径、材积、冠幅分别为4.2 m、5.0 cm、0.0057 m3、2.6 m,变异系数分别为16.01%、20.6%、55.2%、18.39%;极差5.0 m、6.2 cm、0.018 2 m3、3.4 m;与CK2相比,其树高、胸径平均、材积分别提高0.5 m、0.8 cm、0.000 5 m3;增幅达13.5%、19%、9.6%,年均生长量均达1.1 m 、1.2 cm和0.001 9 m3以上,说明在优良林分中开展优良单株选择可获较大增益。其树高生长量变幅为1.9~6.9 m,最大为4号家系,均值为4.8 m;单株树高最高为混合种达6.9 m,其次是4号6.1 m;树高按均值排序为4>CK1>6>12>8>5>2>1>10>7>3>13>9>CK2;胸径生长量变幅为1.5~7.7 cm,最大胸径为6号家系,均值为5.5 cm,单株胸径最大为混合种,为7.7 cm,胸径按均值排序为6>4>7>5>2>1>CK1>12>8>3>13>10>9>CK2;其材积生长量变幅为0.000 3~0.018 5 m3,最大为15(CK1)号家系均值为0.007 m3,单株材积最高为混合种0.018 5 m3,其次是2号0.017 0 m3,材积按均值排序为CK1>6>4>2>5>8>7>1>3>CK2>13>10>9>12;表明米老排母树林经去劣疏伐其遗传品质得到提高,其优树单亲子代差异十分显著。
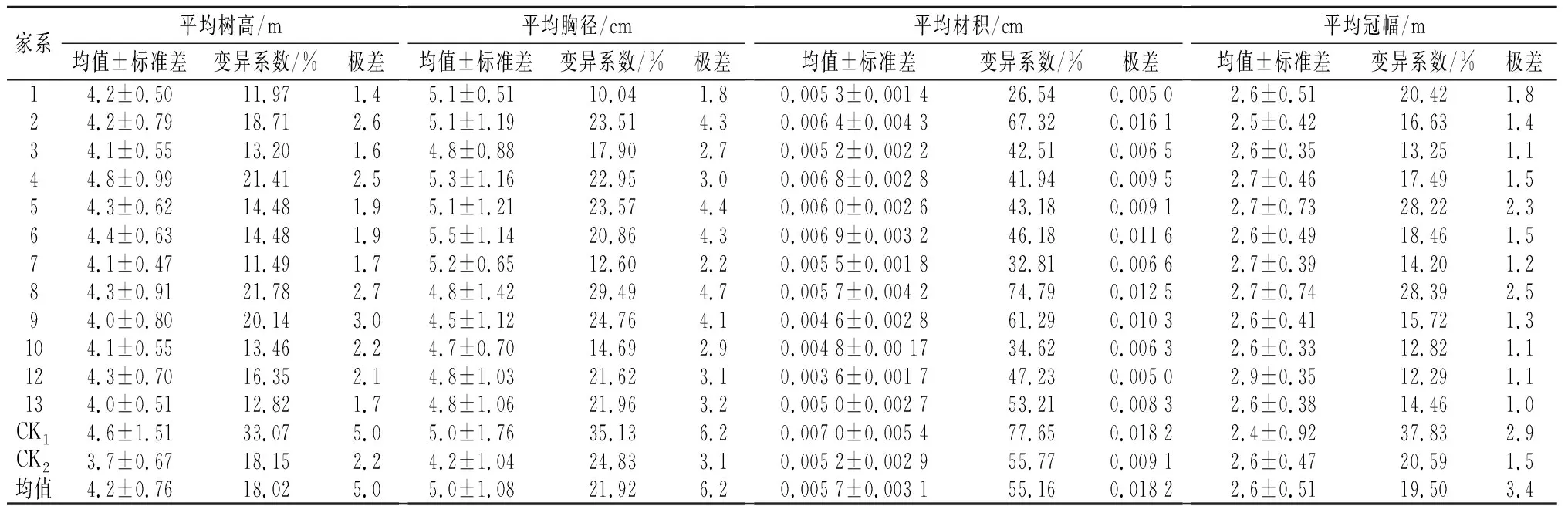
表1 不同家系米老排3年生生长性状指标
2.3 米老排优树单亲子代生长性状方差分析及遗传参数
开展米老排各家系生长性状方差分析,评定各性状家系遗传力的高低是制定遗传改良策略的前提。方差分析可知:米老排优树单亲子代各家系生长性状达极显著差异水平(表2),表明参试子代测定各家系间表型差异趋势明显,这种差异主要是由各家系的遗传基础所决定,开展优良家系选择可获得较大的遗传增益。

表2 米老排优树单亲子代方差分析及遗传力
注:**-极显著差异,*-显著差异
在方差分析基础上估算各性状遗传参数,树高、胸径、材积的家系遗传力分别为0.51、0.56、0.37,与苗期苗高、地径的家系遗传力相比略有下降[2],说明树高、胸径性状受遗传控制较高,选择性状遗传较高的开展子代选择和培育具有重要意义,也为优良家系选择提供依据。
2.4 米老排优树生长量与子代生长相关性分析
开展优树生长量与子代各阶段生长相关性分析,是判断其遗传因子能否独立遗传依据,通过对12株优树的胸径、树高、材积与其子代树高、胸径和苗期地径、苗高相关性分析及相关系数显著性检验(表3),结果表明:优树3年生子代树高与材积,苗高与地径,优树树高、胸径与材积,呈极显著正相关,而子代3年生树高与胸径、胸径与材积,优树3年生子代树高、材积与优树胸径、优树材积呈显著正相关。表明在选择优良家系或者个体中可根据其中一个指标来估测家系生长量。子代3年生树高与苗高、地径、优树树高等其他各性状呈不显著相关。因此开展苗期和子代测定是林木良种选育所必需的[2]。
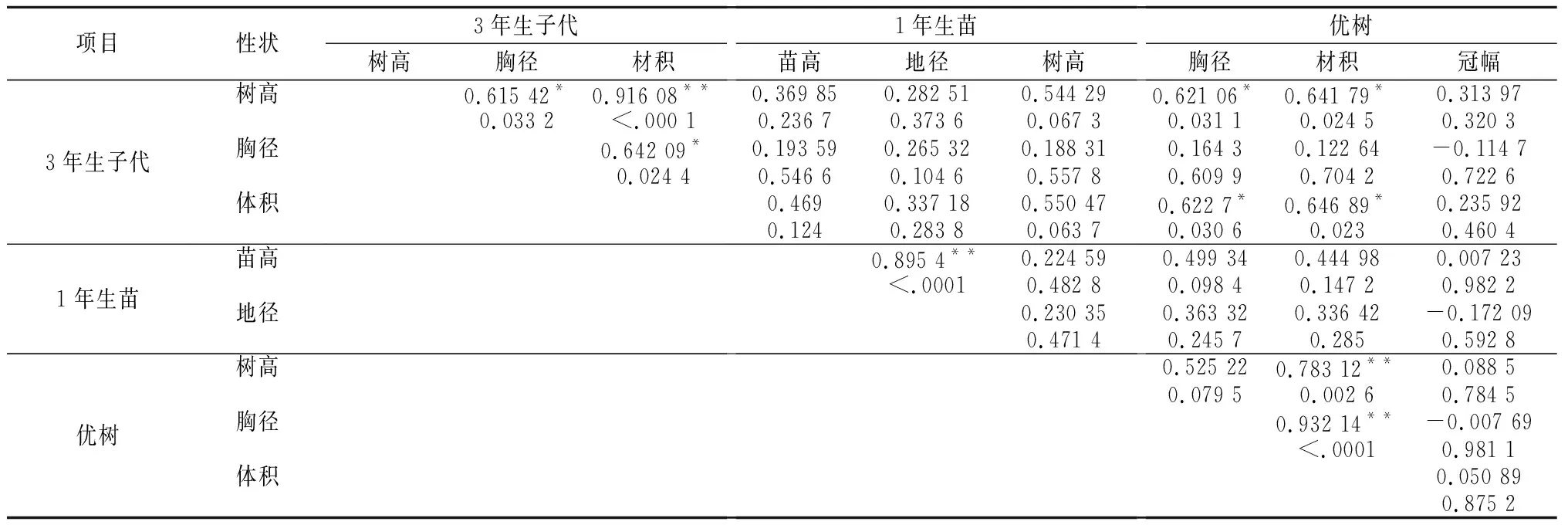
表3 米老排优树与子代生长主要性状相关分析结果
注:r0.01,12=0.651,r0.05,12=0.532
2.5 米老排优良家系和优良个体选择
米老排遗传改良目标是速生丰产,研究表明早期选择是有效的[4],以各性状指标大于群体平均值做为优良家系入选条件,计有2、4、5、6入选。与苗期优良家系相比9、7、3、8淘汰[2]。而2号抗逆性强且速生得以重新入选。以各家系树高、胸径、材积达到或超过群体平均值+1/3标准差为临界值对家系精选[5],仅4、6家系入选。入选家系平均树高、胸径、材积分别为4.6 m、5.4 cm、0.006 9 m3,分别比群体均值高出9.5%、8%、21.1%,比对照CK2高出25.2%、29%、32.7%;遗传增益达5.3%、4.1%、12.1%。在优良家系的基础上进行优良个体的选择,以优良个体树高、胸径、材积达到或超过群体平均值+标准差为标准对单株进行精选,精选出12个优良个体。这些个体分别来源2、4、5、6、8、9、CK1家系,上述家系在苗期选择中多数有入选或家系抗逆性较强[2],这12个优良个体平均树高、胸径为5.6 m、6.8 cm、0.013 3 m3,年均生长量分别达1.8 m、2.3 cm、0.004 4 m3,比米老排人工林生长提高48.1%、263.6%、1770%[6],其现实增益达62.8%、53.3%、56.2%,遗传增益达29.8%、32%、57.8%。这些入选的优良家系、个体性状表现优异,可作为米老排种子园建园材料,其优良个体还可作为无性繁殖的母株加以充分利用[7]。因此,开展优树与优良单株联合选择可获双层次遗传增益[4]。
3 结论
米老排为浅根性阔叶树种,其树叶面积大且浓绿,且髓心空洞,木质部薄,在春季树木未萌动前完成米老排苗木移植是保持苗木水分平衡提高造林成活率关键。
开展优树单亲子代选择是提高林木产量的重要途径,用优树内优良单株的子代造林其增益是家系间和家系内选择增益之和。对12个优树进行子代测定,其优树子代生长量均超过米老排母树林子代生长量。以各性状指标达到或超过群体平均值+1/3标准差为标准,4、6号子代入选为最优家系。入选家系平均树高、胸径、材积为4.6 m、5.4 cm、0.006 9 m3,分别比群体高出9.5%、8%、21.1%,其遗传增益达5.3%、4.1%、12.1%。在优良家系基础上进行优良单株的选择,以优良树高、胸径、材积达到或超过群体平均值+标准差为标准对单株进行精选,精选出12个优良。这12个优良单株平均树高、胸径、材积分别为5.6 m、6.8 cm、0.01 33 m3,其现实增益达62.8%、53.3%、156.2%。遗传增益达29.8%、32%、59.3%。因此,开展优树与优良单株联合选择可获双层次遗传增益。
米老排遗传改良是选择→测定→再选择的循环过程,通过母树林优树单亲本子代测定进行遗传型选择和家系选择,从而筛选出遗传增益高优良家系,可为新建种子园和后续育种提供宝贵的育种材料,从而加快米老排的育种进程。
参考文献:
[1] 洪永辉,林能庆,廖柏林,等.米老排优树选择技术研究[J].林业勘察设计,2015(1):1-4
[2] 洪永辉,林能庆,黄钦忠.米老排优树单亲子代苗期生长变异分析及选择[J].防护林科技,2016,152(5):4-6,34
[3] 黄少伟,谢维辉. 实用SAS编程与林业试验分析[M].广州:华南理工大学出版社,2001:36-44
[4] 洪永辉,林文奖,黄以法,等.马尾松全同胞子代测定及优良单株选择[J].森林与环境学报,2010,30(2):109-114
[5] 贾晨,辜云杰,夏川,等.枫香半同胞家系子代测定及优良家系选择[J].西部林业科学,2014,47(5):122-128
[6] 林能庆,洪永辉,李蔚倩,等.米老排人工林生长规律及生长模型拟合研究[J].林业勘察设计,2017(3): 22-28
[7] 洪永辉,林文奖,黄以法.12年生马尾松种子园半同胞家系生长性状变异分析与优良家系选择[J].南京林业大学学报:自然科学版,2010,34(4):26-30
