长汀侵蚀退化区砍伐迹地马尾松种群动态
2018-06-22杨贤宇李守中宋铁燕王从容吴语嫣宁秋蕊姜良超
杨贤宇,李守中,*,宋铁燕,王从容,刘 聪,吴语嫣,宁秋蕊,姜良超
1 福建师范大学湿润亚热带山地生态国家重点实验室培育基地, 福州 350007 2 福建师范大学地理科学学院, 福州 350007 3 中国科学院植物研究所植被与环境变化国家重点实验室, 北京 100093
在过去的几十年里,人类活动和环境的冲突日益加剧,正以前所未有的速度深刻改变着生态系统的结构和功能。砍伐作为人类对森林生态系统最常见的干扰形式之一,不仅影响森林更新和重建,而且通过影响内部生存环境而引起种群动态变化。研究发现砍伐势必增大植株死亡风险,个体数量的变化改变种群结构,对种群发展和演化产生长期潜在影响[1]。而随着砍伐强度的增大,对林内种群带来的影响也随之改变[2- 4]。轻度择伐能够维持林木良好生长,利于种群结构优化和天然更新[5- 7]。强度择伐则提高种群平均胸径,但会降低种群繁殖能力,致使种群衰退乃至消失[8-9]。同时,砍伐作用因种群的生活史策略和演替阶段不同而不同[10]。演替初期下的砍伐能够降低先锋树种比例,造成演替进程延缓甚至逆行演替。而在顶级群落的砍伐干扰则会阻碍耐阴树种更新,林窗的出现利于先锋树种快速生长[11-12]。目前有关砍伐干扰对植物种群影响已有大量报道[13- 16],但针对干扰后短期内种群响应变化的分析比较薄弱,研究砍伐迹地植物种群动态变化对探究干扰生境下种群生存现状和植被恢复重建具有重要意义。
种群动态是种群生态学研究的核心问题,植物种群变化受控于个体的生命过程(萌发、存活、生长、繁殖及死亡),及影响个体生命过程的生态因子。现阶段我国学者借助年龄分布图、静态生命表和存活分析理论等分析种群动态已取得显著成果[17- 20]。但此类方法多为基于植株数量变化和存活状态得出的定性描述,更多的关注于变化趋势而忽视其内在驱动的重要性。Easterling等[21]将生命率指标纳入考虑提出IPM(积分投影模型,Integral projection model),其从描述性分析和外推预测发展到过程模拟和机制分析[22-23]。IPM一经提出即被众多种群生态学家采用,涉及物种包括伊利里亚大翅蓟(Onopordumillyricum)[22]、黑沙蒿(Artemisiaordosica)[24]、中间锦鸡儿(Caraganaintermedia)[25]、飞廉(Carduusnutans)[26]、类叶升麻(Actaeaspicata)[27]、羊柴(Hedysarumleave)[28]等,而对亚热带退化生态系统中先锋树种—马尾松(Pinusmassoniana)报道则尚未多见[29]。
南方花岗岩山地是我国第二大水土流失区,福建长汀县因环境脆弱性问题突出成为南方水土流失区的典型代表[30-31]。马尾松因其耐贫瘠耐旱,生长周期短等特点成为该地区生态重建的重要建群种,其动态变化指示着亚热带森林群落的演替状态和恢复水平[32]。为了提高林地和森林生产力,砍伐作为重要的营林措施常被采用。同时,随着该地区人口增长和人均耕地面积下降,上山伐薪获取生活能源现象偶有发生[33]。因此探讨砍伐迹地上马尾松先锋种群动态变化及其内在机制,能够为我国南方侵蚀退化区生产营林和植被重建提供一定科学依据。本文通过拟合生命率函数模拟马尾松种群生活史,以期揭示以下问题:马尾松种群在受到砍伐干扰后时期内,(1)种群生命率和大小结构发生何种变化。(2)种群增长率出现上升抑或下降。(3)种群内哪些个体及其何种生命率指标是影响种群增长率变化的关键来源。
1 材料与方法
1.1 研究区概况
长汀县河田镇位于福建省西南部(116°00′45″—116°39′20″E,25°18′40″—26°02′05″N),中亚热带季风性湿润气候,年均降雨量1700mm,多集中于5—7月。年均温18.3℃,最高气温27—30℃(6月),最低气温4—5℃(1月)。地貌以低山、丘陵为主,土壤为花岗岩风化发育的侵蚀红壤,抗蚀性差且酸性强(pH值4—6),属于我国南方面积最大的水土流失区[34]。地带性植被为亚热带常绿阔叶林植被。该地区历史上多次森林滥伐事件导致地表植被群落大面积退化或消失。其后政府于1983年设置水土流失治理试点,经过三十多年的整治,现阶段马尾松纯林和针阔混交林植被以斑块状散布于河田镇周边[35- 36]。
1.2 样地设置与调查
试验地设在长汀县河田镇马坑村,2011年建立马尾松种群动态监测样地,样地面积900m2(30m×30m)。植被以马尾松为建群种,平均基径7.6cm,平均树高2.5m,密度1544株/hm2,少量伴生木荷(Schimasuperba)、黄檀(Dalbergiahupeana)等其他阔叶树种,林下植被主要为铁芒萁(Dicranopterislinearis)。试验地在2012年10月遭受意外砍伐,样地内81%的大个体(基径>50mm)马尾松被砍伐,砍伐数量占马尾松总数19%,砍伐后密度544株/hm2。
2011年7月首次调查时,采用网格状布样方法,将样地划分36个小样方建立方格坐标。对样方内马尾松每木调查并挂牌编号,测量树高>1.3m植株的基径(mm)、胸径(mm)和松果数,测量树高<1.3m植株的基径(mm)和树高(cm)。随后以一年为单位定期重复调查,对新生幼苗挂牌标记并测量地径(mm)和树高(mm)。共获取2013—2016年4期1464株马尾松监测数据。
1.3 数据处理分析
通过预统计分析发现指标基径相对于胸径和树高,能够表达更多的生命率(存活、生长和繁殖)变化信息(Pearson相关系数更高),因此选用基径来描述植株大小并纳入模型构建分析。
1.3.1 IPM积分投影模型
本文采用Esterling等[21]提出的IPM模拟马尾松种群对砍伐干扰的响应变化。其将个体间生长差异纳入模型构建,通过离散时间内连续生长的个体生命率(存活、生长、繁殖等)变化描述种群,模型表达式如下[21]:
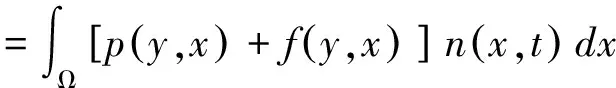
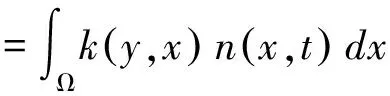
(1)
IPM的核k(y,x)(Kernel)是一个非负曲面,代表从t时刻演变至t+1时刻,植株从个体基径x转变到个体基径y的所有可能性。其由两部分组合而成:p(y,x)=s(x)g(y,x)和f(y,x)=pr(x)fe(x)fd(y)。存活生长函数p(y,x)描述了植株基径为x的个体存活并成长至基径为y的个体的可能性;繁殖函数f(y,x)描述了植株基径为x的个体所产生的基径为y的个体数量。
由于种群内不同生活史阶段马尾松均可能会存活、生长和繁殖新生个体。因此我们将逻辑回归(Logistic regression)用于存活和繁殖概率方程,将成对T检验(Paired Test)用于生长方程。将前1年马尾松的基径(x)和当年存活状态进行回归得到存活方程s(x);将前1年个体基径(x)与当年基径(y)线性回归得到生长方程g(y,x)。将前1年马尾松的基径(x)和当年马尾松的繁殖状态回归得到繁殖概率方程pr(x);将前1年个体基径(x)和当年产生的新生幼苗数回归获得繁殖数量方程fe(x);当年新生幼苗的个体大小分布fd(y)通常用正态分布方程来描述。式中Ω为参与计算的所有个体基径的范围。
1.3.2 种群增长率及弹性分析
运用中点法则(Midpoint rule)[21]将核k(y,x)转化为100阶大转移矩阵K(y,x),其能够输出马尾松种群在调查周期内的种群增长率和关键变化来源等种群信息。转移矩阵的主特征值的生态学意义为种群增长率λ。而弹性分析是预测种群内处于某一阶段的个体的某一生命率的百分比变化对种群增长率λ的影响,可在敏感性分析的基础上通过如下公式计算[24]:
(2)
式中,i为矩阵的行,j为矩阵的列;ij表示矩阵转移因子:即时刻t到时刻t+1的转移率;xij为存活率、正生长率、负生长率或繁殖率。
数据分析处理与模型构建均基于R3.4.1软件平台,此软件为共享统计软件,可在R软件网站(https://www.r-project.org)下载。
2 结果与分析
为了更好的分析种群生活史特征,大致将马尾松大小同生长阶段相对应,基径<10mm的植株个体对应幼苗阶段,基径在10—50mm范围内植株对应幼年树阶段,>50mm马尾松为种群繁殖的主要贡献者—成年树阶段。
2.1 砍伐干扰后生命率变化特征
由图1a,b,c可知,植株的存活概率随着基径增大而上升,但不同大小植株存活概率在调查期内变幅有所差异。2013—2014年马尾松存活状况较好,幼苗和幼年树存活概率高于87%,成年树存活概率高于90%(图1a,表1)。2014—2015年最小基径的存活概率约为75%左右,但最大径级植株存活概率接近100%(图1b,表1)。2015—2016年幼苗植株存活状况出现一定提高,存活概率约为80%左右,但成年树存活状况则相对下降,最大径级的存活概率约为90%(图1c,表1)。
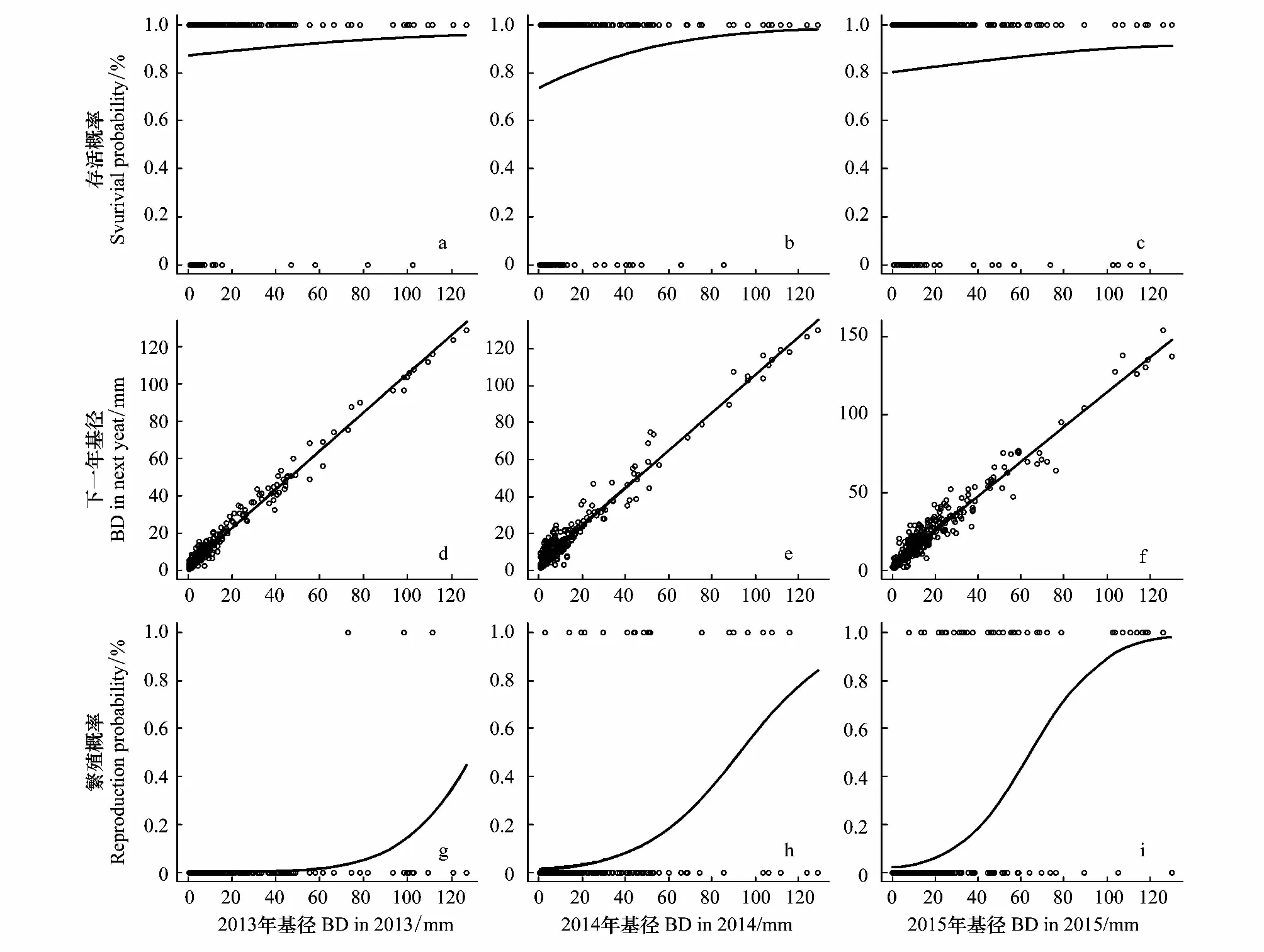
图1 2013—2016年马尾松种群生命率函数Fig.1 Fitting vital rates functions to the Pinus massoniana data during 2013—2016BD:基径 Basal diameter
由图1d,e,f可知,马尾松在调查期内均处于正生长,表现为生长函数斜率大于1。种群植株平均生长量从2013—2014年的(2.47±0.18)mm上升至2015—2016年的(6.39±0.37)mm,其中幼苗生长量从(1.77±0.15)mm上升至(4.38±0.38)mm,幼年树生长量从(3.74±0.42)mm上升至(7.13±0.50)mm,成年树生长量从(4.20±1.30)mm上升至(11.26±2.22)mm。另一方面,负增长的个体比例约为种群总数的10%—16%。
由图1g,h,i可知,马尾松的繁殖概率随着植株基径增大而上升。在调查期内,大径级马尾松(基径>100mm)的繁殖概率得到了显著提升,其从高于15%逐步提升至高于90%(表1)。种群内处于繁殖状态的马尾松从3株发展至38株,松果总数从110颗增长至334颗。但每年新生幼苗数量在调查期内从68株下降至25株(表1)。
表1用于构建2013—2016年(时期1—3)马尾松种群IPM核的统计模型和参数估计
Table1StatisticalmodelsandparameterestimatesusedtoconstructthekernelfortheIntegralProjectionModelsofPinusmassonianaduring2013—2016(period1—3)

种群参数Demographic parameters时期Period函数Model 种群参数Demographic parameters时期Period函数Model 存活概率sSurvival probability/%1Logit(s)=1.920(0.206)+0.009(0.010)xn=340,R2=0.006,P<0.053μ=4.455(0.443)+1.109(0.016)xn=302,R2=0.943,P<0.0012Logit(s)=1.026(0.154)+0.023(0.009)xn=339,R2=0.035,P<0.05生长残差σ2Variance of 1σ2=5.157(1.188)+0.251(0.044)xn=290,R2=0.099,P<0.0013Logit(s)=1.400(0.170)+0.008(0.007)xn=386,R2=0.006,P<0.05growth/mm2σ2=17.510(3.046)+0.321(0.109)xn=268,R2=0.028,P<0.01繁殖概率prReproduction probability/%1Logit(PR)=-7.498(1.753)+0.057(0.018)xn=340,R2=0.450,P<0.013σ2=16.761(4.352)+0.967(0.153)xn=302,R2=0.115,P<0.0012Logit(s)=-4.267(0.400)+0.046(0.007)xn=339,R2=0.030,P<0.001新生苗大小分布Size distribution of 1正态分布均值=2.184方差=2.303,n=683Logit(s)=-3.913(0.348)+0.061(0.009)xn=386,R2=0.426,P<0.001new recruits/mm2正态分布均值=1.553方差=3.624,n=59下一年的个体大小μFuture size/mm1μ=1.937(0.213)+1.035(0.008)xn=290,R2=0.987,P<0.0013正态分布均值=2.056方差=2.484,n=252μ=4.221(0.350)+1.019(0.012)xn=268,R2=0.961,P<0.001
模型构建以个体基径(mm)为自变量,括号内数值为估计参数的标准误差,n为用于方程统计的植物个体数,R2为拟合优度,P<0.01表示方程的参数通过显著性检验。
2.2 砍伐干扰后种群增长率变化和弹性分析

图2 砍伐迹地马尾松种群结构变化特征(2014—2016) Fig.2 Population structural variation characteristics of Pinus massoniana in logged forestland (2014—2016)
由图2可知,砍伐迹地上马尾松植株尺寸集中于小径级范围,幼苗和幼树的数量比例很高,达到90%左右。总株数在调查期内由341株增长至387株,最大植株径级从基径130mm左右生长至基径170mm左右。幼年树和成年树数量增加,比例分别为75.8%和50%,而幼苗数量从224株减少至113株,减少比例约50%。
由表2可知,种群增长率在调查期内均小于1,且呈继续减小趋势,表明这一时期马尾松种群处于衰退态势,且衰退程度不断加深。基于植株生命率的弹性分析显示(图3),马尾松的存活弹性值明显高于其他生命率的弹性值,表明植株的存活对砍伐后的种群发展至关重要。小径级植株的生长有利于种群发展,大径级植株的生长不利于种群发展,表现为小径级植株生长的弹性值为正值,而大径级植株则为负值。在调查周期内,基径40—60mm的马尾松生长的弹性值从正值转变为负值。而植株负增长和繁殖的弹性值在不同生活史阶段上均接近于零,表明对种群发展的贡献程度十分有限。基径在0—5mm左右的幼苗存活和生长的弹性值处于种群内最高水平,显示这一径级范围的马尾松对整个种群重要性最大。
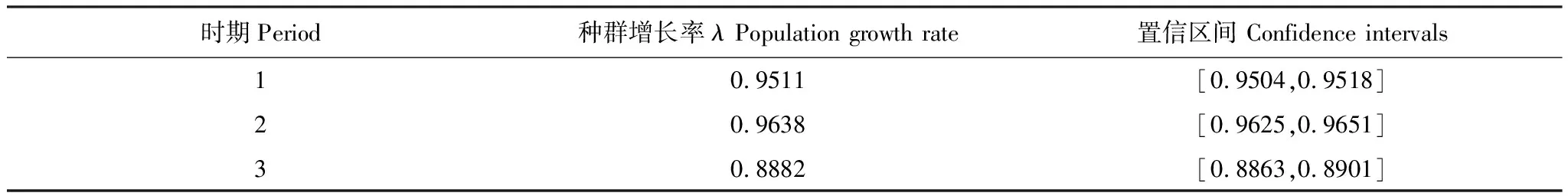
表2 不同时期马尾松种群增长率及置信区间比较
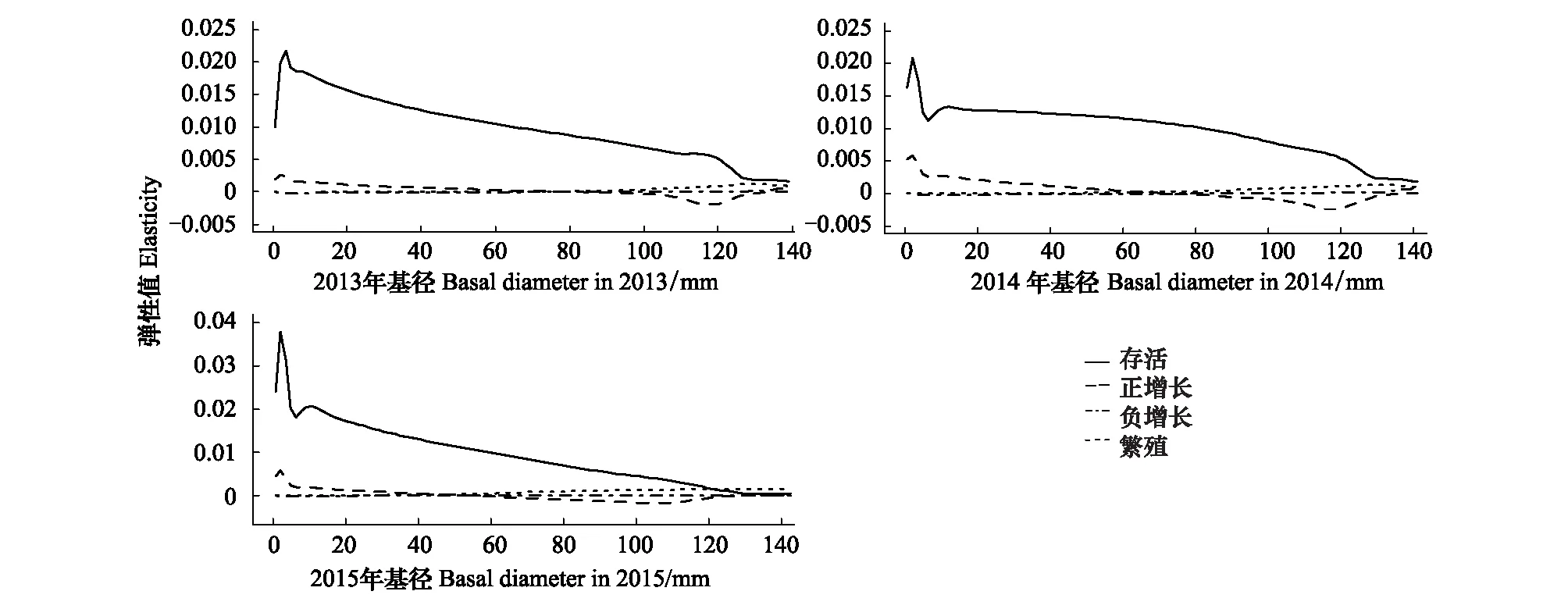
图3 2013—2016年马尾松生活史弹性分析Fig.3 Elasticity analysis of life stage for the integral projection model fitted to Pinus massoniana during 2013—2016
3 讨论
砍伐迹地上,马尾松种群不同生活史阶段个体及生命率随时间均不断发生变化。(1)就存活生命率而言:幼苗个体在生理生态特征上发育并不成熟,因此抵抗外界环境变化能力较弱[37]。由于样地内部分地块裸露,夏季水热变化剧烈,马尾松幼苗在生境胁迫下难以度过幼年期,小个体马尾松是种群死亡的主要来源。同时,砍伐作业后土壤养分、理化性质等发生变化,幼苗存活产生年际波动变化[13,38]。大个体死亡因素主要来自砍伐作业中树干折损降低了植株的后续光合能力[38- 39]。相对于幼苗和幼年树,成树抵御环境变化能力较强,因此成活率年际变化较小。(2)就生长生命率而言:马尾松作为强向阳性树种,光资源相对于其他生境资源,对个体存活和生长尤为重要[11]。对成年树来说,林分密度的降低扩大了其营养空间,更大的林冠使得植株得到更好的生长。同时林下光照条件的改善减轻了遮光作用,幼苗和幼年树生长也得到一定加速[2,41]。但是,对于部分枝叶损失严重的个体,光合产物下降导致暂时的生长发育滞缓。脱皮作为马尾松一种生物特性,当直径生长速度低于树皮脱落速度时,砍伐迹地上植株出现负增长现象。(3)就繁殖生命率而言:砍伐迹地上马尾松繁殖概率随着时间逐渐提升,表明在个体资源内部分配上,生殖配置比重逐渐增加。虽然马尾松每年产果,但由于成年树在种群内比例较低,加之铁笀萁对种子进入土壤及幼苗萌发的机械阻碍作用,新生苗数相对种群来说仍然十分稀少且逐年下降。马尾松作为中国南方亚热带森林演替中的先锋树种,其依靠大量的种子产生和幼苗建成弥补高死亡率,来实现种群扩张和更新[29]。可以预见,当成年树个体衰老死亡后,该种群也将因低比例的幼苗更新而逐渐灭亡,被其他树种取代。
马尾松作为演替进程中的先锋种,其种群常常处于不稳定状态。虽然已有研究发现一定程度的择伐有利于种群自我更新和维持[40- 41],但研究发现:当81%的大个体植株(基径>50mm)被去除后,砍伐迹地上马尾松种群处于衰退状态,且衰退程度不断加深。弹性分析表明,此时个体的存活是对种群最为重要的生命率,同其他干扰生境下弹性分析结果一致[24-25,44]。小径级植株的存活弹性值显著高于其他生活史阶段,且随着时间延长,其相对重要性逐渐加强,因此在砍伐迹地上应首要保证幼苗、幼树的存活以维持种群。加之生长弹性值在该阶段为正值,因此可选取长势良好的幼苗、幼树,底部挖施肥穴,定期追肥,促进其生长以加速种群生物量积累。另一方面马尾松作为先锋树种,其扩张依靠大量的种子繁殖弥补个体的高损失率。虽然弹性值显示繁殖对种群贡献不大,但砍伐带来的新生苗的巨大下降也会潜在的导致种群衰退。因此可适当的人为播撒马尾松种子和移植新生苗以促进砍伐迹地上马尾松种群恢复。研究砍伐对先锋植物种群动态变化的影响,有助于深入理解干扰生境下种群响应变化过程,为南方侵蚀退化地马尾松林的管理和保护提供一定依据。
4 结论
(1)在长汀侵蚀退化区砍伐迹地上,马尾松种群在伐后4年内幼苗数量大幅度减少,幼年树和成树数量上升。死亡在生活史各个阶段均有出现,幼苗是死亡的主要来源,其存活率年际波动较其他阶段更为明显。生长量和生长增量随着植株增大和时间增加而上升。个体繁殖概率逐年提升,但新生苗数量持续下降。
(2)经过砍伐后(81%的大径级植株被伐),马尾松种群处于衰退状态(λ<1),且衰退程度不断加深。这一时期,所有保留木的存活对种群维持至关重要,小径级植株的正生长利于种群增长,大径级植株的正生长不利于种群增长,而负生长和繁殖对种群增长率的影响均十分有限。
(3)在生态恢复过程中,种群内生物因素和外部环境驱动因素既有规律性,又有随机性,马尾松作为先锋种,其种群动态过程的复杂性使得种群增长率在某些年份可能会更高或更低。因此在未来应将长时间尺度下的环境波动和气候变化等因素纳入考虑,完善研究结果。
参考文献(References):
[1] Grogan J, Landis R M, Free C M, Schulze M D, Lentini M, Ashton M S. Big-leaf mahoganySwieteniamacrophyllapopulation dynamics and implications for sustainable management. Journal of Applied Ecology, 2014, 51(3): 664- 674.
[2] 曾冀, 雷渊才, 贾宏炎, 蔡道雄, 唐继新. 桂西南马尾松人工林生长对不同强度采伐的动态响应. 林业科学研究, 2017, 30(2): 335- 341.
[3] 周梦丽, 张青, 亢新刚, 杨英军, 王全军. 云冷杉天然林乔木树种组成及物种多样性对择伐强度的动态响应. 植物科学学报, 2016, 34(1): 56- 66.
[4] 包也, 于大炮, 周莉, 周旺明, 孟莹莹, 吴志军, 郭焱, 王晓雨, 代力民. 择伐对阔叶红松林资源可持续利用的影响. 生态学报, 2015, 35(1): 31- 37.
[5] 张悦, 易雪梅, 王远遐, 姬兰柱, 吴培莉. 采伐对红松种群结构与动态的影响. 生态学报, 2015, 35(1): 38- 45.
[6] 杨梅. 不同强度人为干扰对马尾松生长的影响. 福建林业科技, 2004, 31(2): 68- 71.
[7] 林思祖, 杨梅, 曹子林, 刘洪波, 陈艳. 不同强度人为干扰对马尾松地上部分生物量及生产力的影响. 西北植物学报, 2004, 24(3): 516- 522.
[8] 姚克平. 马尾松人工林不同抚育间伐强度的生长效应研究. 湖北林业科技, 2006, (3): 10- 12.
[9] 方萍. 间伐对马尾松中龄林生长的影响. 福建林业, 2017, (2): 38- 41, 37- 37.
[10] 齐麟, 代力民, 于大炮, 周莉, 钟磊, 苏东凯. 采伐对长白山阔叶红松林乔木群落的影响. 湖南农业科学, 2009, (8): 129- 132.
[11] 丁圣彦, 宋永昌. 常绿阔叶林演替过程中马尾松消退的原因. 植物学报, 1998, 40(8): 755- 760.
[12] 刘峰, 谭畅, 王红, 龙江平. 基于LiDAR的亚热带次生林林窗对幼树更新影响分析. 农业机械学报, 2017, 48(3): 198- 204.
[13] 邱仁辉, 周新年, 杨玉盛. 森林采伐作业的环境影响及其保护对策. 中国生态农业学报, 2003, 11(1): 130- 132.
[14] 简耘, 石磊, 李丹, 张纯淳, 石苗苗, 王嵘, 陈小勇. 森林砍伐对苦槠种群遗传结构的影响. 生态学报, 2008, 28(12): 6228- 6234.
[15] 郑丽凤. 闽北天然次生林择伐强度的阈值和择伐策略研究[D]. 福州: 福建农林大学, 2008.
[16] 郑丽凤, 周新年. 择伐强度对天然林树种组成及物种多样性影响动态. 山地学报, 2008, 26(6): 699- 706.
[17] 陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214- 217.
[18] 于大炮, 周莉, 董百丽, 代力民, 王庆礼. 长白山北坡岳桦种群结构及动态分析. 生态学杂志, 2004, 23(5): 30- 34.
[19] 李金昕, 吴定军, 章世鹏, 何显湘, 陈坚, 史清茂, 胥晓. 四川米仓山自然保护区台湾水青冈种群生命表及动态分析. 植物研究, 2016, 36(1): 68- 74.
[20] 陈倩, 杨晔, 史琛媛, 徐学华. 木兰围场天然次生林优势种群的结构与动态. 应用生态学报, 2017, 28(1): 37- 46.
[21] Easterling M R, Ellner S P, Dixon P M. Size-specific sensitivity: applying a new structured population model. Ecology, 2000, 81(3): 694- 708.
[22] Ellner S P, Rees M. Integral projection models for species with complex demography. The American Naturalist, 2006, 167(3): 410- 428.
[23] Zuidema P A, Jongejans E, Chien P D, During H J, Schieving F. Integral projection models for trees: a new parameterization method and a validation of model output. Journal of Ecology, 2010, 98(2): 345- 355.
[24] Li S L, Yu F H, Werger M J A, Dong M, Zuidema P A. Habitat-specific demography across dune fixation stages in a semi-arid sandland: understanding the expansion, stabilization and decline of a dominant shrub. Journal of Ecology, 2011, 99(2): 610- 620.
[25] Li S L, Yu F H, Werger M J A, Dong M, Ramula S, Zuidema P A. Understanding the effects of a new grazing policy: the impact of seasonal grazing on shrub demography in the Inner Mongolian steppe. Journal of Applied Ecology, 2013, 50(6): 1377- 1386.
[26] Jongejans E, Allen M, Shea K. Spatial dynamics of invasiveCarduusthistles//Marinova C, ed. 19th International Congress on Modelling and Simulation. Perth: Am Nat, 2011: 2514- 2520.
[27] Dahlgren J P, Ehrlén J. Linking environmental variation to population dynamics of a forest herb. Journal of Ecology, 2009, 97(4): 666- 674.
[28] Li S L, Yu F H, Werger M J A, Dong M, During H J, Zuidema P A. Mobile dune fixation by a fast-growing clonal plant: a full life-cycle analysis. Scientific Reports, 2015, 5: 8935.
[29] 林培治. 福建长汀水土流失区马尾松种群动态及其机制研究[D]. 福州: 福建师范大学, 2013.
[30] Zhu H J. Strategies on Eco-restoration in the Subtropical Mountain Ecosystem Fragility Areas, China: Based on the Achievement of Eleven Years′ Research in Changting County. Journal of Natural Resources, 2015, 6(3): 139- 145.
[31] 岳辉. 强度侵蚀山地不同治理措施对植被生长的影响及其生态效益分析. 亚热带水土保持, 2008, 20(3): 23- 27.
[32] 彭绍云. 长汀县水土流失区树种结构现状与调整. 亚热带水土保持, 2005, 17(1): 29- 30, 53- 53.
[33] 马志阳, 查轩. 南方红壤区侵蚀退化马尾松林地生态恢复研究. 水土保持研究, 2008, 15(3): 188- 93.
[34] 陈志彪. 花岗岩侵蚀山地生态重建及其生态环境效应[D]. 福州: 福建师范大学, 2005.
[35] 范小明. 福建水蚀荒漠化地区种草治理效果与地表植被演替规律. 现代农业科技, 2014, (2): 273- 274.
[36] 姚雄, 余坤勇, 刘健, 杨素萍, 何平, 邓洋波, 俞欣妍, 陈樟昊. 南方水土流失严重区的生态脆弱性时空演变. 应用生态学报, 2016, 27(3): 735- 745.
[37] 韩文萍, 丁贵杰, 鲍斌. 不同种源马尾松对干旱胁迫的生理生态响应. 中南林业科技大学学报, 2012, 32(5): 25- 29.
[38] 邱仁辉, 周新年, 杨玉盛. 择伐对林地土壤物理性质影响及作业技术. 福建林学院学报, 2001, 21(4): 301- 303.
[39] 邱仁辉, 周新年. 不同强度的择伐作业对保留木与幼树幼苗的影响. 森林工程, 1997, 13(3): 5- 7.
[40] Figueira A M E S, Miller S D, De Sousa C A D, Menton M C, Maia A R, da Rocha H R, Goulden M L. Effects of selective logging on tropical forest tree growth. Journal of Geophysical Research, 2008, 113(G1): 243- 250.
[41] 黄承标, 马承彪, 曹继钊, 许涛, 陶大燕, 莫兰英, 牙小红, 黄燕妮. 不同立地及间伐强度对马尾松人工幼林生长的影响. 西北林学院学报, 2013, 28(1): 141- 145.
[42] 黄小波, 李帅锋, 苏建荣, 苏磊. 择伐对思茅松自然种群结构和空间分布格局的影响. 生态学报, 2015, 35(24): 8241- 8250.
[43] 李瑞霞, 闵建刚, 彭婷婷, 刘娜, 郝俊鹏, 王东, 关庆伟. 间伐对马尾松人工林植被物种多样性的影响. 西北农林科技大学学报: 自然科学版, 2013, 41(3): 61- 68.
[44] He D, Wang Q G, Franklin S B, Jiang M X. Transient and asymptotic demographics of the riparian speciesEupteleapleiospermumin the Shennongjia area, central China. Biological Conservation, 2013, 161: 193- 202.
