黄麻纤维产量与主要农艺性状的相关分析
2018-06-15张列梅祁建民方书生张力岚2方平平张立武
徐 益 张列梅 祁建民 苏 梅 方书生,2 张力岚,2方平平 张立武,2
黄麻纤维产量与主要农艺性状的相关分析
徐 益1,2,**张列梅1,**祁建民1苏 梅1方书生1,2张力岚1,2方平平1张立武1,2,*
1福建农林大学作物遗传育种与综合利用教育部重点实验室 / 福建省作物设计育种重点实验室/作物科学学院, 福建福州 350002;2福建农林大学海峡联合研究院基因组与生物技术中心, 福建福州 350002
研究黄麻纤维产量与主要农艺性状的相关性, 可为高产育种与生产利用提供科学依据。本文159份不同来源黄麻种质资源的12个农艺性状对纤维产量即单株干皮重的影响表明, 各性状的变异系数变化在11.89%至38.50%之间, 表现出丰富的遗传变异。黄麻纤维产量与各性状均呈极显著正相关, 其中, 与单株鲜皮重、株高、始花期的相关系数较大, 分别为0.814、0.760和0.648。黄麻纤维产量和单株鲜皮重、株高、出麻率、鲜皮厚的回归方程达显著水平, 其标准回归系数依次为0.443、0.437、0.291和0.113。通径分析显示, 单株鲜皮重、株高在决定黄麻纤维产量时起主要作用。出麻率的相关系数(0.253)与直接通径系数(0.291)表现基本一致, 说明出麻率直接对黄麻纤维产量起作用, 具极显著正相关。因此, 在黄麻高产育种中, 应该以始花期、单株鲜皮重、株高、出麻率与鲜皮厚为主要筛选对象, 兼顾综合性状的改良。
黄麻; 农艺性状; 纤维产量; 相关分析; 通径分析
黄麻(spp)是重要的韧皮部纤维作物之一, 有2个栽培种, 即长果种黄麻()和圆果种黄麻()[1-3], 染色体数均为2= 14。目前, 黄麻主要在温带、亚热带和热带地区种植, 分布在印度、孟加拉国、中国、乌兹别克斯坦、尼泊尔、越南、缅甸、津巴布韦、泰国、埃及等国家。由于黄麻韧皮部纤维具有可生物降解、吸湿性能好和散湿水分快等许多优良特性, 被广泛应用于纺织、造纸、建筑与医药等领域[4-5]。
黄麻以收获茎皮为目的, 其纤维产量受很多农艺性状影响, 如株高、分枝高、鲜皮厚、节数、分枝数等[1]。各个性状皆具有数量性状的遗传特点, 且相互关联, 单单改良某一个性状难以使纤维产量大幅增长。因此, 为了更好地提高黄麻的育种和生产效率, 十分有必要基于其农艺性状进行相关及通径分析。在黄麻农艺性状的研究中, 主要从农艺性状鉴定及遗传力、遗传相关等方面分析[6-8]。祁建民等[6]鉴定不同黄麻品种的主要数量性状遗传力, 认为株高的广义遗传力和狭义遗传力较高, 对其进行早代选择可靠性较大; 孙家曾等[7]仅利用12份黄麻种质资源材料分析株高与单株干皮产量等经济性状的相关性, 认为单株干皮产量与株高性状极显著正相关; 张加强等[8]利用主成分分析和聚类分析的方法对26个黄麻种质资源产量性状研究表明, 前3个主因子提供了原始性状88.247%的生物信息, 并在此基础上用欧氏距离聚类图将26个黄麻种质分成四大类。在育种过程中发现, 黄麻纤维产量虽与各农艺性状有着密切的联系, 且不同的农艺性状之间呈现出不同程度的相关性, 然而哪个性状对黄麻纤维产量有着最密切的联系, 应该以哪个性状作为最主要的筛选对象?这些问题并不清楚。因此, 黄麻纤维产量与主要农艺性状的相关性分析及通径分析对于黄麻育种与生产利用有着十分重要的意义。
本研究一方面扩大黄麻种质资源的数量, 另一方面增加农艺性状的数量, 利用159份不同来源的黄麻种质资源, 先对各农艺性状之间进行相关分析, 进而对纤维产量和不同农艺性状进行回归分析, 再估算通径系数, 旨在分析黄麻不同农艺性状之间的关系以及不同农艺性状对黄麻纤维产量的影响, 为黄麻的高产育种与生产利用提供理论依据。
1 材料与方法
1.1 供试材料
159份不同来源的黄麻种质资源[9]均由福建农林大学麻类遗传育种与综合利用实验室提供。
1.2 试验设计
分别于2014年4月30日和2016年5月1日在福建省三明市尤溪县福建农林大学洋中科教基地农场, 按2行区, 行长3.50 m, 株行距1.20 m × 0.10 m, 采用单因素随机区组设计种植供试材料, 边行种植两行为保护行, 田间管理同大田。
1.3 农艺性状考察
参考《黄麻种质资源描述规范和数据标准》[10]考察黄麻主要农艺性状(附表1)。试验初始数据中,株高(1)、分枝高(2)、皮厚(3)、单株鲜茎重(4)、单株鲜皮重(5)、节数(6)、分枝数(7)等均来自黄麻工艺成熟时采集的10株左右样本考种所得的平均值, 出麻率(8)是根据(单株干皮重/单株鲜茎重)计算而来。
1.4 数据统计分析
用SPSS 20.0处理数据, 利用两年数据的平均值分析各生育期性状与单株干皮重()的相关系数。生育期性状包括现蕾期、始花期、工艺成熟期与种子成熟期。进行黄麻单株干皮重与8个经济性状(1~8)以及各性状之间的相关分析。再以单株干皮重()为因变量, 进行回归分析并计算通径系数, 来分析不同农艺性状对黄麻纤维产量的直接或间接影响[11]。
2 结果与分析
2.1 黄麻农艺性状的表现
从表1可以看出, 159份黄麻种质资源群体各性状的变异系数变化在11.89%至38.50%之间, 最大的是出麻率, 最小的是种子成熟期。表现出丰富的遗传变异, 属于随机种质资源群体, 适合后续的相关与通径分析。
2.2 黄麻纤维产量与生育期的相关分析
从表2可以看出, 生育期各个阶段与黄麻的纤维产量均表现出极显著的正相关, 且相关系数极为相似, 变化在0.584至0.648之间, 说明黄麻整个生育期越长, 黄麻纤维产量越高。始花期对单株干皮重的相关系数最大(0.648), 反映出始花期对黄麻纤维产量的作用较明显。
表1 黄麻主要农艺性状的统计分析
Table 1 Statistical analysis of main agronomic traits in jute
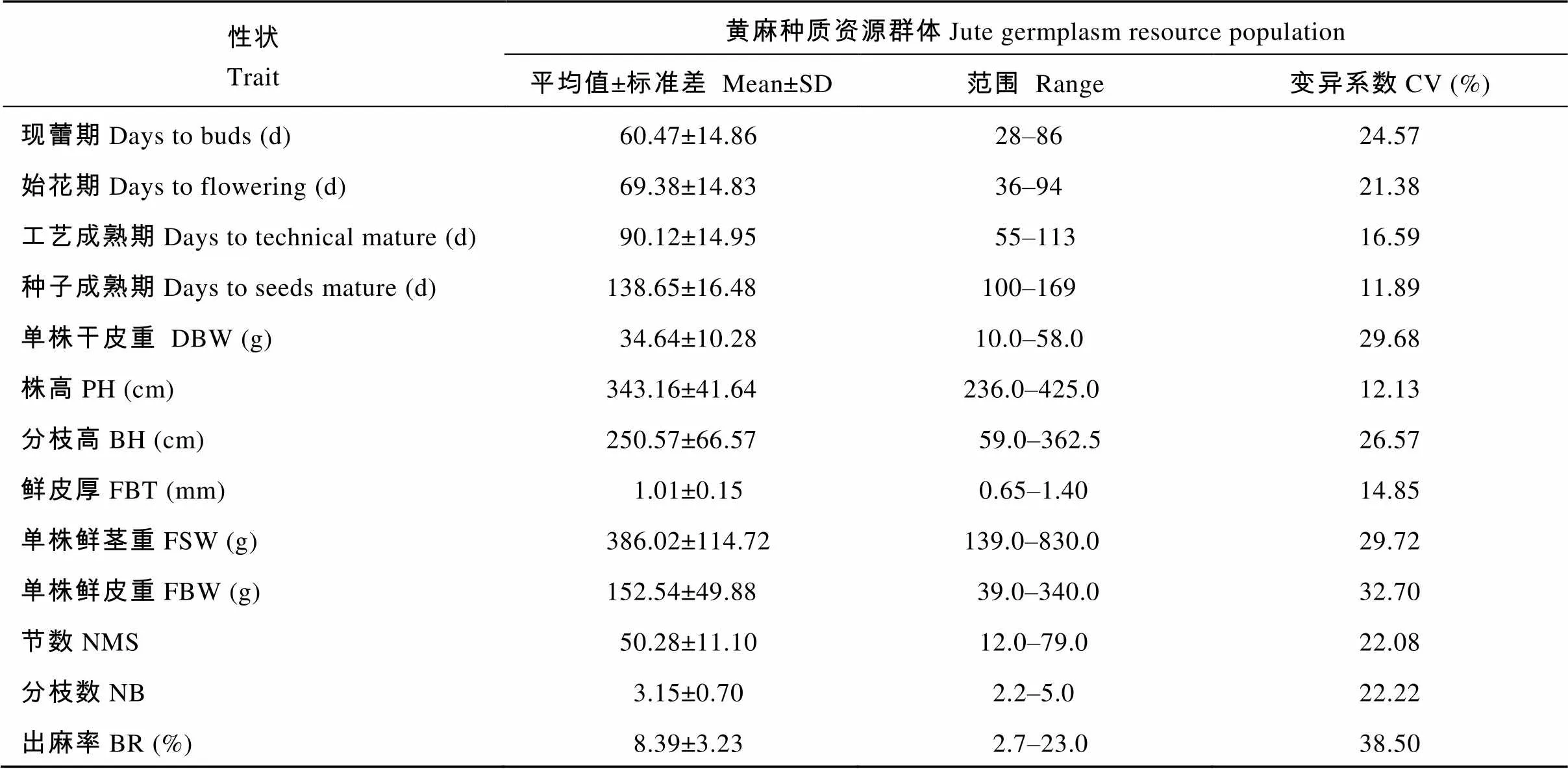
性状Trait黄麻种质资源群体Jute germplasm resource population 平均值±标准差 Mean±SD范围 Range变异系数CV (%) 现蕾期Days to buds (d)60.47±14.8628–8624.57 始花期Days to flowering (d)69.38±14.8336–9421.38 工艺成熟期Days to technical mature (d)90.12±14.9555–11316.59 种子成熟期Days to seeds mature (d)138.65±16.48100–16911.89 单株干皮重 DBW (g)34.64±10.2810.0–58.029.68 株高PH (cm)343.16±41.64236.0–425.012.13 分枝高BH (cm)250.57±66.5759.0–362.526.57 鲜皮厚FBT (mm)1.01±0.150.65–1.4014.85 单株鲜茎重FSW (g)386.02±114.72139.0–830.029.72 单株鲜皮重FBW (g)152.54±49.8839.0–340.032.70 节数NMS50.28±11.1012.0–79.022.08 分枝数NB3.15±0.702.2–5.022.22 出麻率BR (%)8.39±3.232.7–23.038.50
DBW: dry bark weight per plant; PH: plant height; BH: branching height; FBT: fresh bark thickness; FSW: fresh stem weight per plant; FBW: fresh bark weight per plant; NMS: nodes of main stem; NB: number of branches; BR: bark rate; SD: standard deviation; CV: coefficient of variation.

表2 黄麻纤维产量与各个生育期性状的相关分析
**表示在<0.01水平显著。**means significance at<0.01. DBW: dry bark weight per plant.
2.3 黄麻纤维产量与经济性状的相关分析
从表3可以看出, 除出麻率与分枝数、单株鲜皮重、鲜皮厚, 分枝数与鲜皮厚、节数, 鲜皮厚与株高和分枝高之外, 黄麻的不同经济性状之间均存在不同程度相关。株高、分枝高、鲜皮厚、单株鲜茎重、单株鲜皮重、节数、分枝数、出麻率8个经济性状皆与纤维产量极显著相关, 其中相关系数由大到小依次为单株鲜皮重(0.814)>株高(0.760)>单株鲜茎重(0.743)>分枝高(0.673)>节数(0.658)>鲜皮厚(0.402)>分枝数(0.337)>出麻率(0.253)。黄麻出麻率虽然与纤维产量极显著正相关, 但与单株鲜茎重却显著负相关(–0.237), 这与出麻率的计算公式有关。
2.4 黄麻纤维产量与经济性状的回归分析
从表4可以看出, 单株鲜皮重对单株干皮重的影响最大, 其标准回归系数为0.443, 其后依次为株高0.437、出麻率0.291、鲜皮厚0.113。因此, 在黄麻的高产育种中, 单株鲜皮重、株高、出麻率、鲜皮厚相对于另外4个经济性状对纤维产量具有更直接的作用。但不能忽略各经济性状之间相互影响的间接作用。
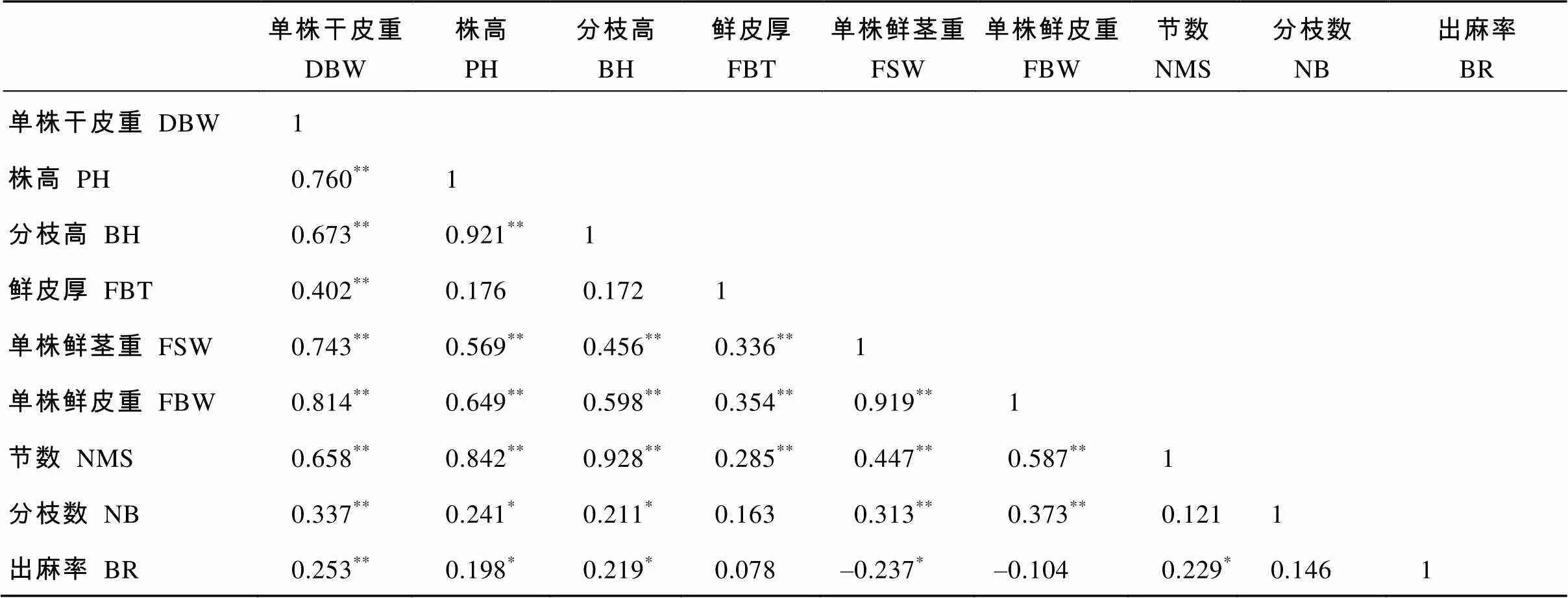
表3 黄麻纤维产量和8个经济性状的相关分析
*和**分别表示在<0.05和<0.01水平显著。缩写同表1。
*and**mean significance at<0.05 and<0.01, respectively. The abbreviations are the same as those given in Table 1.
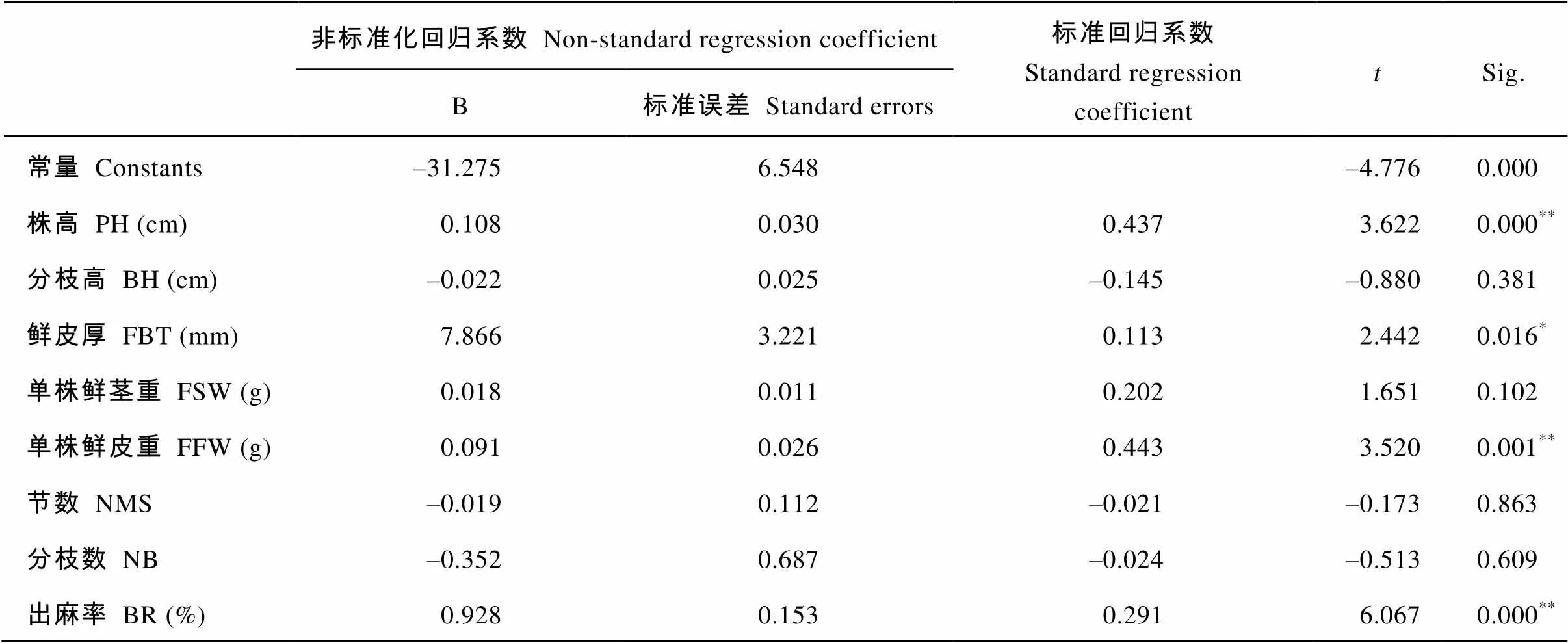
表4 黄麻纤维产量和经济性状的多元标准回归分析
*和**分别表示在<0.05和<0.01水平显著。缩写同表1。
*and**mean significance at<0.05 and<0.01, respectively. The abbreviations are the same as those given in Table 1.
2.5 黄麻纤维产量与经济性状的通径分析
株高对单株干皮重的相关系数1y=0.760, 株高和分枝高之间的相关系数为12=0.921 (表3); 分枝高对黄麻干皮重的标准回归系数即直接通径系数为2y= –0.145 (表4), 利用这些数据可以计算出株高(1), 分枝高2)对单株干皮重影响的间接通径系数为2y×12= –0.145×0.921= –0.134。同理可求出株高(1)对鲜皮厚(3)的间接通径系数以及所有变量间的间接通径系数。最后将数据归纳为表5。
8个经济性状与单株干皮重极显著相关。在此基础上, 通过比较这5个性状的直接通径系数(即标准回归系数)发现, 株高对单株干皮重的直接通径系数为0.437, 单株鲜皮重的直接通径系数为0.443, 远远大于其他性状, 说明单株鲜皮重和株高对纤维产量尤为重要。而像分枝高、单株鲜茎重、节数, 与单株干皮重呈现出较高的相关性, 但其与黄麻单株干皮重的直接通径系数分别为–0.145、0.202和–0.021, 其中, 分枝高和节数与单株干皮重的直接通径系数甚至表现为负值。结合其与单株干皮重的间接通径系数可知, 分枝高、单株鲜茎重、节数对黄麻干皮重的影响大部分都是通过影响株高和单株鲜皮重的间接作用而产生的。
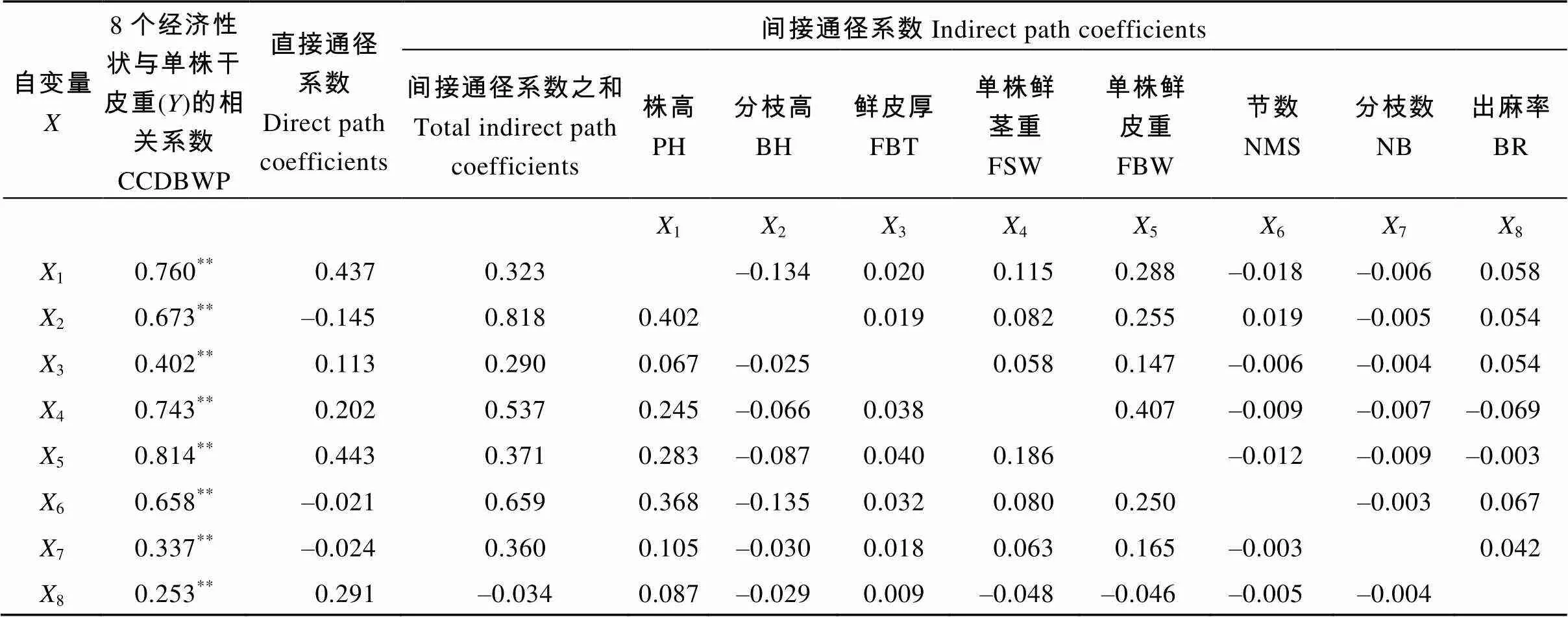
表5 黄麻纤维产量与8个经济性状通径分析
1: 株高;2: 分枝高;3: 鲜皮厚;4: 单株鲜茎重;5: 单株鲜皮重;6: 节数;7: 分支数;8: 出麻率。**表示在< 0.01水平显著。缩写同表1。
1: plant height;2: branching height;3: fresh bark thickness;4: fresh stem weight per plant;5: fresh bark weight per plant;6: nodes of main stem;7: number of branches;8: bark rate.**means significance at< 0.01. The abbreviations are the same as those given in Table 1. CCDBWP: correlation coefficients of dry bark weight per plant () with eight economic traits.
此外, 在8个经济性状中, 出麻率与黄麻单株干皮重的相关系数最小, 但其相关系数与直接通径系数表现基本一致。反映出出麻率对黄麻产量影响可以排除其他性状的作用, 直接对黄麻纤维产量产生作用, 且呈现显著正相关。因此出麻率对黄麻的高产起着至关重要的作用。
3 讨论
深入研究种质资源的遗传变异特征, 有利于扩大种质资源的创新与利用, 明确育种目标[9, 12]。本研究表明, 参试材料的13个农艺性状的变异系数变化范围较广, 说明在这159份种质资源中这些农艺性状存在较大差异, 可为黄麻高产育种提供了材料基础。各农艺性状的变异系数依次为出麻率>单株鲜皮重>单株鲜茎重>单株干皮重>分枝高>现蕾期>分枝数>节数>始花期>工艺成熟期>鲜皮厚>株高>种子成熟期。其中, 种子成熟期、株高、鲜皮厚、工艺成熟期等性状变异系数均小于20%, 始花期、节数、分枝数、现蕾期、分枝高、单株干皮重、单株鲜茎重变异系数在20%~30%之间, 而单株鲜皮重与出麻率是复合性状, 变异系数均大于30%, 受环境影响较大。这与张加强等[13]的研究结果基本一致, 但本研究试验材料更多, 代表性更强。
相关系数能说明各农艺性状的相关程度[14-17]。12个农艺性状都与单株干皮重呈极显著相关, 暗示纤维产量与这些性状间可能存在一因多效[18]或紧密连锁的QTL (quantitative trait loci)族[19-20], 这方面的研究有待深入。在各个生育期性状中, 始花期对黄麻纤维产量的作用较明显。在8个经济性状中, 相关系数较大的有单株鲜皮重(0.814)、株高(0.760)、单株鲜茎重(0.743)、分枝高(0.673)、节数(0.658)、鲜皮厚(0.402)等。郑云雨等[15]利用4个品种分析发现, 株高、鲜皮厚、单株鲜茎重与单株干皮重显著相关, 但认为分枝高、出麻率与单株干皮重关系不大, 这与本研究结果不完全一致。出麻率作为干皮重与鲜茎重的比值, 应该与干皮重正相关, 与鲜茎重负相关, 本研究体现这一点。在单株鲜皮重、株高、鲜皮厚基础上, 以出麻率来评价纤维产量会比单纯以单株鲜皮重、株高、鲜皮厚、单株鲜茎重更客观。因此, 在筛选黄麻高产种质时, 除了把单株鲜皮重、株高和鲜皮厚作为主要目标性状外, 还应着重提高出麻率。
黄麻高产育种中, 既要通过相关分析来剖析各农艺性状对纤维产量的作用, 也要通过通径分析来剖析各农艺性状之间的相互关系。目前, 通径分析广泛应用于资源筛选、品种比较、自交系选育、品种资源等众多领域的综合评估[15-17]。由相关分析可以得知, 本研究涉及的12个农艺性状均与单株干皮重呈极显著相关。通径分析发现, 分枝高、单株鲜茎重、节数、分枝数等农艺性状都是通过株高和单株鲜皮重间接作用于黄麻的单株干皮重, 才呈现显著的相关性。这表明, 只进行相关分析, 并不能正确解释各性状对黄麻纤维产量的影响, 必须结合后面的通径分析, 重新梳理各性状对产量的实质影响, 以及各性状之间的相互联系。尽管黄麻单株鲜皮重、株高、单株鲜茎重、分枝高、节数、鲜皮厚等经济性状与纤维产量(单株干皮重)皆呈现显著相关, 但直接通径系数反映出仅单株鲜皮重、株高、出麻率、鲜皮厚与纤维产量的回归方程达显著水平, 其中单株鲜皮重、株高对黄麻单株干皮重的直接作用最大。因此, 在育种实践中, 选择具有更重的单株鲜皮重、较高的株高和较大鲜皮厚的品系是黄麻高产育种最为切实可靠的途径。对于出麻率, 其相关系数与直接通径系数大小差不多, 反映出与黄麻干皮产量之间的相关几乎不受其他农艺性状的影响, 所以尽量提高黄麻的出麻率也是高产育种中极为重要的一环。至于其他的经济性状, 其与黄麻单株干皮重的极显著相关, 大多数都是通过单株鲜皮重和株高表现出来的。因此, 对于黄麻高产育种来说, 在适宜的生育期基础上, 应优先考虑单株鲜皮重、株高、鲜皮厚以及出麻率。当然, 本文仅针对12个农艺性状与纤维产量做了相关与通径分析, 目的在于为黄麻的高产育种提供相应的理论依据。在育种实践中, 还应考察综合性状的改良, 如纤维品质[21]、抗逆性[22]等, 从优质稳产等角度进一步研究和探讨。
4 结论
不同来源黄麻种质资源的生育期、株高、分枝高、鲜皮厚、单株鲜茎重、单株鲜皮重、节数、分枝数、出麻率均与纤维产量(单株干皮重)表现出极显著正相关。黄麻纤维产量与单株鲜皮重、株高、出麻率、鲜皮厚的回归方程达显著水平, 其中单株鲜皮重与株高在决定黄麻的干皮重时起主导作用, 而出麻率的相关系数与直接通径系数大小相当, 体现出直接对纤维产量产生作用。在高产育种及生产中, 在适宜的生育期、单株鲜皮重、株高与鲜皮厚得到保证的前提下, 应尽量提高黄麻的出麻率。
[1] 熊和平. 麻类作物育种学. 北京: 中国农业科学技术出版社, 2008. pp 208–296Xiong H P. Breeding Sciences of Bast and Leaf Fiber Crops. Beijing: China Agricultural Science and Technology Press, 2008. pp 208–296 (in Chinese)
[2] 张加强, 陈常理, 骆霞红, 金关荣. 中国育成黄麻主要品种间的亲缘关系分析. 中国农业科学, 2015, 48: 4008–4020Zhang J Q, Chen C L, Luo X H, Jin G R. Analysis of the coefficient of parentage among major jute cultivars in China., 2015, 48: 4008–4020 (in Chinese with English abstract)
[3] Islam M S, Saito J A, Emdad E M, Ahmed B, Islam M M, Halim A, Hossen Q M, Hossain M Z, Ahmed R, Hossain M S, Kabir S M, Khan M S, Khan M M, Hasan R, Aktar N, Honi U, Islam R, Rashid M M, Wan X, Hou S, Haque T, Azam M S, Moosa M M, Elias S M, Hasan A M, Mahmood N, Shafiuddin M, Shahid S, Shommu N S, Jahan S, Roy S, Chowdhury A, Akhand A I, Nisho G M, Uddin K S, Rabeya T, Hoque S M, Snigdha A R, Mortoza S, Matin S A, Islam M K, Lashkar M Z, Zaman M, Yuryev A, Uddin MK, Rahman M S, Haque M S, Alam M M, Khan H, Alam M. Comparative genomics of two jute species and insight into fibre biogenesis., 2017, 3: 16223
[4] Rana M K, Arora K, Singh S, Singh A K. Multi-locus DNA fingerprinting and genetic diversity in jute (spp.) based on sequence-related amplified polymorphism., 2012, 22: 1–8
[5] Mir R R, Rustgi S, Sharma S, Singh R, Goyal A, Kumar J, Gaur A, Tyagi A, Khan H, Sinha M K, Balyan H S, Gupta P K. A preliminary genetic analysis of fibre traits and the use of new genomic SSRs for genetic diversity in jute., 2007, 161: 413–427
[6] 祁建民, 卢浩然, 郑云雨, 王英娇. 黄麻数量性状遗传关系分析. 作物学报, 1991, 17: 145–150 Qi J M, Lu H R, Zheng Y Y, Wang Y J. Genetic relationship analysis of quantitative traits in jute., 1991, 17: 145–150 (in Chinese with English abstract)
[7] 孙家曾, 余隆其, 何广文. 黄麻主要数量性状遗传力和相关性的研究. 中国农业科学, 1981, 14(3): 25–32 Sun J Z, Yu L Q, He G W. Heritability and correlation studies of major quantitative traits in jute., 1981, 14(3): 25–32 (in Chinese with English abstract)
[8] 张加强, 陈常理, 骆霞红, 金关荣. 26份黄麻种质资源产量性状的主成分聚类分析极其评价. 植物遗传资源学报, 2016, 17: 475–482 Zhang J Q, Chen C L, Luo X H, Jin G R. Evaluation of yield components of 26 jute germplasm resources by principal component cluster analysis., 2016, 17: 475–482 (in Chinese with English abstract)
[9] Zhang L, Yuan M, Tao A, Xu J, Lin L, Fang P, Qi J. Genetic structure and relationship analysis of an association population in jute (spp.) evaluated by SSR markers., 2015, 10: e0128195
[10] 粟建光. 黄麻种质资源描述规范和数据标准. 北京: 中国农业出版社, 2005. pp 7–27Su J G. Descriptors and Data Standard for Jute (L. &L.). Beijing: China Agriculture Press, 2005. pp 7–27 (in Chinese)
[11] 杜家菊, 陈志伟. 使用SPSS线性回归实现通径分析的方法. 生物学通报, 2010, 45(2): 4–6 Du J J, Chen Z W. Method of path analysis using SPSS linear regression., 2010, 45(2): 4–6 (in Chinese with English abstract)
[12] Zhang L, Li A, Wang X, Xu J, Zhang G, Su J, Qi J, Guan C. Genetic diversity of kenaf () evaluated by inter-simple sequence repeat (ISSR)., 2013, 51: 800–810
[13] 张加强, 骆霞虹, 陈常理, 朱关林, 金关荣. 圆果种黄麻主要经济性状与纤维产量的相关及灰色关联分析. 中国麻业科学, 2015, (2): 70–74 Zhang J Q, Luo X H, Chen C L, Zhu G L, Jin G R. Correlation and gray relational analysis on main economic characters and fiber yield of white jute., 2015, (2): 70–74 (in Chinese with English abstract)
[14] 卢浩然, 郑云雨, 朱秀英, 王英娇. 黄麻七个经济性状遗传力的研究. 中国麻作, 1980, (1): 6–8 Lu H R, Zheng Y Y, Zhu X Y, Wang Y J. Genetic studies of seven economic traits in jute., 1980, (1): 6–8 (in Chinese with English abstract)
[15] 郭安平, 龚友才. 圆果种黄麻主要农艺性状的遗传相关及通径分析. 中国麻作, 1988, (2): 6–9 Guo A P, Gong Y C. Genetic correlation and path analysis of the main agronomic traits in white jute., 1988, (2): 6–9(in Chinese with English abstract)
[16] 郑云雨, 卢浩然, 王英娇, 祁建民. 黄麻主要经济性状相关的研究. 中国麻作, 1985, (3) : 38–41 Zheng Y Y, Lu H R, Wang Y J, Qi J M. Study on the correlation of main economic characters of jute.1985, (3): 38–41 (in Chinese with English abstract)
[17] 郭安平, 龚友才. 长果种黄麻品种主要农艺性状与单株产量关系的分析. 中国麻作, 1988, (4): 13–18 Guo A P, Gong Y C. Analysis of relationship between main agronomic characters and yield per plant of long fruit jute varieties.1988, (4): 13–18 (in Chinese with English abstract)
[18] Xue W, Xing Y, Weng X, Zhao Y, Tang W, Wang L, Zhou H, Yu S, Xu C, Li X, Zhang Q. Natural variation inis an important regulator of heading date and yield potential in rice., 2008, 40: 761–767
[19] Zhang L W, Yang G S, Liu P W, Hong D F, Li S P, He Q B. Genetic and correlation analysis of silique-traits inL. by quantitative trait locus mapping., 2011, 122: 21–31
[20] Shi J, Li R, Qiu D, Jiang C, Long Y, Morgan C, Bancroft I, Zhao J, Meng J. Unraveling the complex trait of crop yield with quantitative trait loci mapping in., 2009, 182: 851–861
[21] 郑云雨, 祁建民, 李维明, 缪小红, 王英娇, 林培青, 卢浩然. 黄麻产量和纤维品质性状典型相关与双重筛选逐步回归分析. 福建农林大学学报(自然科学版), 1994, (1): 17–20 Zheng Y Y, Qi J M, Li W M, Miu X H, Wang Y J, Lin P Q, Lu H R. Canonical correlation and double screening stepwise regression analysis ofyield and fiber quality traits in jute.(Nat Sci Edn), 1994, (1): 17–20 (in Chinese with English abstract)
[22] 卢瑞克, 杨泽茂, 戴志刚, 许英, 唐蜻, 程超华, 陈基权, 粟建光. 50份长果黄麻种质资源耐盐性鉴定评价. 植物遗传资源学报, 2017, 18(6): 1–11.Lu R K, Yang Z M, Dai Z G, Xu Y, Tang Q, Cheng C H, Chen J Q, Su J G. Evaluation for salt tolerance of 50 jute germplasm resources (L.)., 2017, 18(6): 1–11 (in Chinese with English abstract)
附表1 黄麻主要农艺性状考察方法
Supplementary table 1 Evaluation methods of main agronomic traits in jute

农艺性状 Agronomic traits考察方法 Methods of evaluation 生育期Growth stages现蕾期Days to buds当小区麻株开始现蕾(直径约2 mm, 肉眼可见)后, 隔1 d一次, 上午9:00–10:00观测, 记录现蕾株数。以试验小区全部麻株为观测对象, 50%的植株现蕾的日期为现蕾期。表示方法为“年月日”, 格式为“YYYYMMDD”。如“20160629”, 表示2016年06月29日现蕾。 始花期Days to flowering当小区开第1朵花后, 隔1 d一次, 上午9:00–10:00观测, 记录开花株数。以试验小区全部麻株为观测对象, 50%的植株开花的日期为开花期。表示方法和格式同现蕾期。 工艺成熟期Days to technical mature当植株出现上花下果(长果种花多果少, 圆果种果多花少)后, 表明已达到工艺成熟期。以试验小区全部麻株为观测对象, 记录小区2/3以上植株达到工艺成熟的日期为工艺成熟期。表示方法和格式同现蕾期。 种子成熟期Days to seeds mature当植株2/3以上的蒴果变成褐色时, 表明黄麻已经入种子成熟期。以试验小区全部麻株为观测对象, 记录小区2/3以上植株达到种子成熟的日期为种子成熟期。表示方法和格式同现蕾期。 节数Nodes of main stem在黄麻植株的结果期, 蒴果完全成熟前3~5 d, 从试验小区中部随机取样10株为观测对象, 调查每株从茎秆子叶节至第1个有效分分枝节位的节数。单位为节, 精确到1节。 分枝数Number of branches以调查节数时采集的样株为观测对象, 调查每株的一级有效分枝数。单位为个, 精确到1个。 分枝高Branching height以调查节数时采集的样株为观测对象, 度量每株从茎秆基部到第一有效分枝节位的距离。单位为cm, 精确到0.1 cm。 株高Plant height在黄麻植株的工艺成熟期, 从试验小区中部随机取样10株为观测对象, 度量每株从茎秆最基部到主茎生长点的距离。单位为cm, 精确到0.1 cm。
(续附表1)

农艺性状 Agronomic traits考察方法 Methods of evaluation 鲜皮厚Fresh bark thickness以度量株高时采集的样株为观测对象, 用螺旋测微器, 又名千分卡尺(精度为1/10000)度量每株从茎秆基部以上全株高度1/3处的鲜麻皮厚度。单位为mm, 精确到0.01 mm。 单株鲜茎重Fresh stem weight per plant黄麻鲜茎指除根去叶后的鲜茎秆。在黄麻植株的工艺成熟期, 从试验小区中部随机取样10株, 用1/100的电子天平称取鲜茎重量, 再换算成单株鲜茎重。单位为g, 精确到0.1 g。 单株鲜皮重Fresh bark weight per plant黄麻鲜皮指鲜茎上剥下的新鲜麻皮。在黄麻植株的工艺成熟期, 从试验小区中部随机取样10株, 用1/100的电子天平称取鲜皮重量, 再换算成单株鲜皮重。单位为g, 精确到0.1 g。 单株干皮重Dry bark weight per plant黄麻干皮指鲜茎上剥下后, 完全晒干的麻皮。在黄麻植株的工艺成熟期, 从试验小区中部随机取样10株, 用1/100的电子天平称取干皮重量, 再换算成单株干皮重。单位为g, 精确到0.1 g。
Correlation Analysis between Yield of Bast Fiber and Main Agronomic Traits in Jute (spp.)
XU Yi1,2,**, ZHANG Lie-Mei1,**, QI Jian-Min1, SU Mei1, FANG Shu-Sheng1,2, ZHANG Li-Lan1,2, FANG Ping-Ping1, and ZHANG Li-Wu1,2,*
1Key Laboratory for Genetics, Breeding and Multiple Utilization of Crops of Ministry of Education / Fujian Key Laboratory for Crop Breeding by Design / College of Crop Science, Fuzhou 350002, Fujian, China;2Center for Genomics and Biotechnology of Haixia Institute of Science and Technology, Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian, China
Studies on correlation between yield of bast fiber and main agronomic traits will provide scientific evidence to the breeding for high yield and reproduction of cultivars in jute. In this study, we analyzed the impact of 12 agronomic traits from 159 different jute germplasm resources on bast fiber yield (dry bark weight per plant). The variation coefficients of different traits varied from 11.89% to 38.50%, indicating abundant genetic variation among these traits. Bast fiber yield had significantly positive correlations with other traits, among which the relative high correlation coefficients between fiber yield and fresh bark weight per plant, plant height, days to flowering were 0.814, 0.760, and 0.648 respectively. The regression equation between bast fiber yield and fresh bark weight per plant, plant height, fresh bark thickness, bark rate was significant, with standard regression values of 0.443, 0.437, 0.291, and 0.113 respectively. Path coefficient analysis indicated that fresh bark weight per plant and plant height played a leading role in the determination of bast fiber yield. Also, the correlation coefficient (0.253) and standard regression coefficient (0.291) of bark rate were very close, showing that bark rate directly affects bast fiber yield, with a significant positive correlation between bast fiber yield and bark rate. Therefore, in breeding jute varieties with high bast fiber yield, it is imperative to take days to flowering, fresh bark weight per plant, plant height, bark rate and fresh bark thickness as main selection criteria with consideration for the improvement of comprehensive traits improvement.
jute; agronomic traits; yield of bast fiber; correlation analysis; path coefficient analysis
本研究由国家自然科学基金项目(31771369), 福建省科技厅对外合作项目(2015I001), 国家现代农业产业技术体系建设专项(CARS-19-E06), 农业部东南黄红麻实验观测站(农科教发2011)和福建省麻类种质资源共享平台(2010N2002)资助。
This study was supported by the National Natural Science Foundation of China (31771369), the Foreign Cooperation Project of Science and Technology Department in Fujian, China (2015I001), the China Agriculture Research System (CARS-19-E06), the Experiment Station of Jute and Kenaf in Southeast China (Nongkejiaofa 2011), and the Construction of Germplasm Resources Platform for Bast Fiber Crops in Fujian, China (2010N2002).
URL:http://kns.cnki.net/kcms/detail/11.1809.S.20180126.1315.022.html
2017-10-09;
2018-01-08;
2018-01-26.
10.3724/SP.J.1006.2018.00859
张立武, E-mail: lwzhang@fafu.edu.cn, zhang_liwu@hotmail.com, Tel: 0591-87644898
**同等贡献(Contributed equally to this work)
徐益, E-mail: 1275924118@qq.com; 张列梅, E-mail: 398166832@qq.com
