不同来源木聚糖酶及其组合对肉鸡肠道黏膜形态与二糖酶活性及其基因表达的影响
2018-06-14黄婧溪李忠良臧旭鹏余小华左建军冯定远
黄婧溪 李忠良* 臧旭鹏 余小华 张 涛 左建军** 冯定远**
(1.华南农业大学动物科学学院,广州 510642;2.赢创德固赛(中国)投资有限公司,北京 100600)
麦类饲粮中含有木聚糖等非淀粉多糖(non-starch polysaccharides,NSP),其抗营养功能一直受到人们的关注。在动物饲粮中添加木聚糖酶等非淀粉多糖酶,可破坏植物细胞壁,提高饲粮中淀粉和蛋白质等营养物质的利用率[1];降低消化道食糜黏度,减少疾病的发生[2];减缓非淀粉多糖对动物肠道黏膜的损伤,改善黏膜形态[3]。此外,麦类饲粮中含有大量的淀粉,其部分水解产物包括麦芽糖、麦芽三糖和极限糊精等不能直接被肠道吸收,必须由黏膜二糖酶水解成单糖后才能被吸收利用[4]。而饲粮中的碳水化合物含量直接影响小肠黏膜二糖酶水解反应,使二糖酶具有很强的适应性。
目前有关单一木聚糖酶和含有木聚糖酶的复合酶对肉鸡肠道黏膜形态影响的研究得到的结论[5-6]不尽相同,其原因是影响酶制剂作用效果的因素很多,从而表现不同的生产效应。针对该问题,冯定远[7]提出了组合酶的概念,所谓组合酶是指利用酶催化的协同作用,在催化水解同一底物的不同来源和特性的酶中选择具有互补性的2种或2种以上酶配合而成的酶制剂。
综上,本试验拟研究不同来源木聚糖酶及其组合对肉鸡肠道黏膜形态与二糖酶活性及其基因表达的影响,从而探讨组合型木聚糖酶与肉鸡肠道黏膜形态及碳水化合物消化的关系。
1 材料与方法
1.1 试验动物与试验设计
将1 560只1日龄且体重[(38±2) g]相近的健康新兴黄羽肉公鸡随机分为4组,每组6个重复,每个重复65只。Ⅰ组(对照组)饲喂低能量低蛋白质的小麦-豆粕型基础饲粮,试验组(Ⅱ、Ⅲ和Ⅳ组)分别饲喂在基础饲粮中添加4 000 U/kg的黑曲霉来源单一木聚糖酶1#、康氏木霉来源单一木聚糖酶2#和组合型木聚糖酶(木聚糖酶1#∶木聚糖酶2#=1∶1)。基础饲粮组成及营养水平见表1。试验期63 d。试验期间肉仔鸡自由采食,充足饮水。
1.2 样品采集
分别于21、42、63日龄,在每组的每个重复随机选取2只肉鸡,颈动脉放血屠宰,宰前不禁食。分别采取十二指肠中段、空肠中段组织样品。用磷酸盐缓冲液(PBS)缓慢的冲洗掉食糜,剪成1 cm左右的小段置于固定液中固定。然后沿纵向剖开肠道,用4 ℃预冷的PBS冲洗,吸水纸吸干。分别刮取十二指肠中段、空肠中段黏膜样品置液氮速冻,-80 ℃冷冻保存备用。
1.3 检测指标
1.3.1 肠道黏膜形态测定
制作肠道石蜡切片,经苏木精-伊红染色后,参照Frankel等[8]的方法用显微镜和形态学图像分析系统测量肠道黏膜绒毛高度(villous height,VH)及与之相连的隐窝深度(crypt depth,CD)、绒毛宽度(villous width,VW),并计算绒毛高度/隐窝深度(V/C)值。各指标测定3次,取平均值作为测定数据。
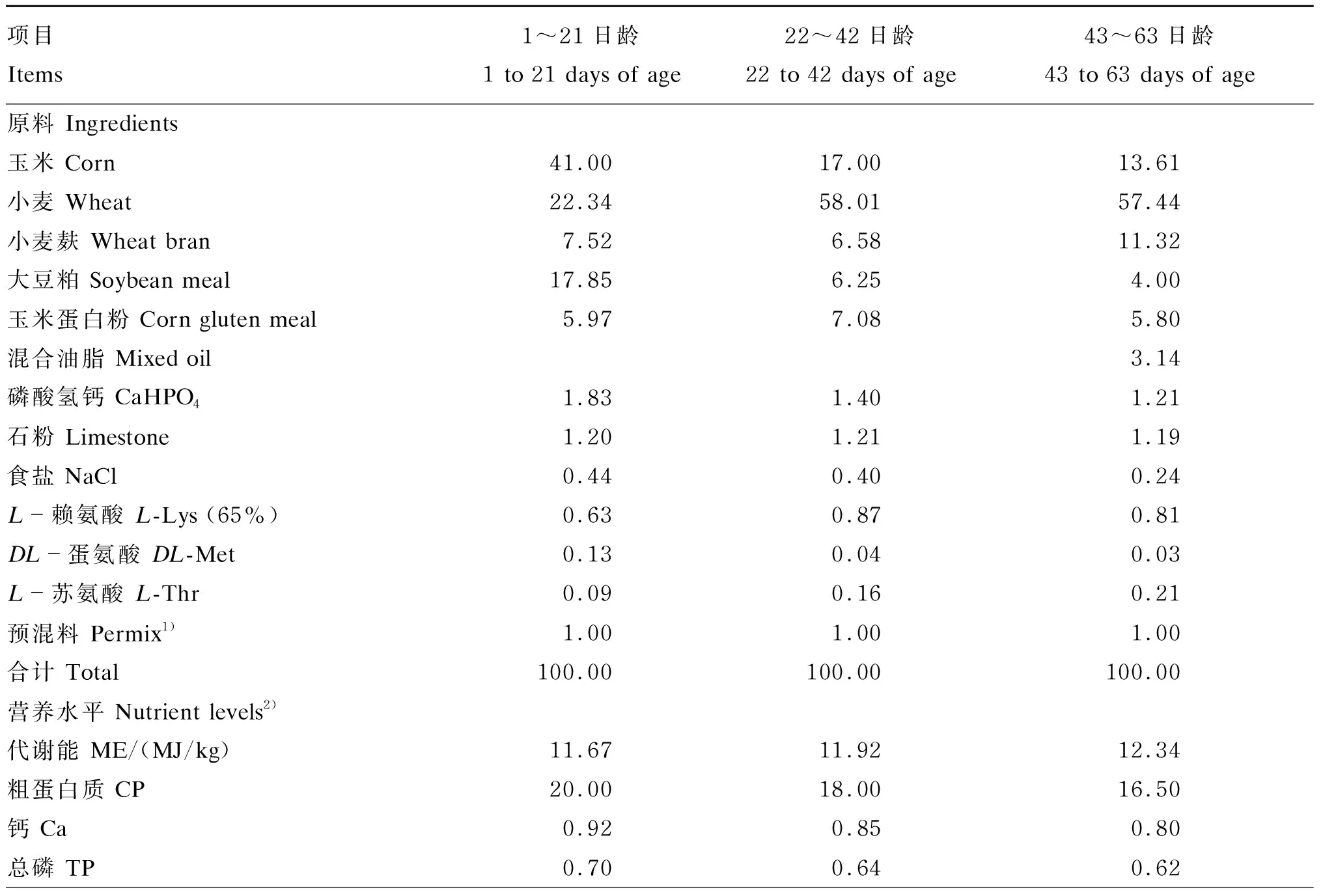
表1 基础饲粮组成及营养水平(风干基础)

续表1项目 Items1~21日龄1 to 21 days of age22~42日龄22 to 42 days of age43~63日龄43 to 63 days of age有效磷 AP0.450.410.38赖氨酸 Lys1.090.940.85蛋氨酸 Met0.580.470.40蛋氨酸+半胱氨酸 Met+Cys0.910.770.67苏氨酸 Thr0.780.690.68色氨酸Trp0.210.180.16
1)1~21日龄、22~42日龄和43~63日龄的预混料分别使用温氏食品集团黄羽肉小鸡、肉中鸡和肉大鸡预混料The premixes of 1 to 21, 22 to 42 and 43 to 63 days of age used the premixes of yellow-feather chickens, young broilers and adult broilers of Wens Food Group, respectively。每千克黄羽肉小鸡预混料提供Provided the following per kg of premix of yellow-feathered chickens:蛋氨酸 Met 120 g,Cu 1 000 mg,Fe 8 000 mg,Mn 8 000 mg,Zn 5 000 mg,I 35 mg,Se 25 mg,VA 1 500 000 IU,VD 300 000 IU,VE 4 000 mg,VK 300 mg,VB1300 mg,VB6700 mg,VB123 mg,胆碱 choline 61 000 mg,叶酸 folic acid 150 mg,烟酸 niacin 5 000 mg,泛酸 pantothenic acid 2 000 mg,生物素 biotin 15 mg,促生长剂 growth promoting agent 500 mg,防霉剂 anti-mildew agent 2 000 mg;每千克黄羽肉中鸡预混料提供Provided the following per kg of premix of yellow-feather young broilers:蛋氨酸 Met 110 g,Cu 880 mg,Fe 5 500 mg,Mn 6 600 mg,Zn 4 400 mg,I 38.5 mg,Se 16.5 mg,VA 1 320 000 IU,VD 330 000 IU,VE 3 300 mg,VK 220 mg,VB1220 mg,VB6550 mg,VB123.3 mg,胆碱 choline 56 000 mg,叶酸 folic acid 110 mg,烟酸 niacin 4 400 mg,泛酸 pantothenic acid 1 650 mg,生物素 biotin 13.2 mg,防霉剂 anti-mildew agent 2 000 mg;每千克黄羽肉大鸡预混料提供Provided the following per kg of premix of yellow-feather adult broilers:蛋氨酸 Met 80 g,Cu 800 mg,Fe 5 000 mg,Mn 6 000 mg,Zn 4 000 mg,I 35 mg,Se 25 mg,VA 1 000 000 IU,VD 200 000 IU,VE 3 000 mg,VK 200 mg,VB1200 mg,VB6500 mg,VB122 mg,胆碱 choline 41 000 mg,叶酸 folic acid 50 mg,烟酸 niacin 3 500 mg,泛酸 pantothenic acid 1 500 mg,生物素 biotin 10 mg,防霉剂 anti-mildew agent 2 000 mg。
2)营养水平均为计算值。All nutrient levels were measured values.
绒毛高度:从绒毛顶端至隐窝开口处的垂直距离。
绒毛宽度:绒毛最宽处的距离。
隐窝深度:从隐窝开口至隐窝基部的垂直距离。
1.3.2 肠道黏膜生化指标测定
称取0.5 g左右的肠道黏膜,加入4 mL 0.1 mol/L pH为6.8的顺丁烯二酸缓冲液,冰浴匀浆,4 ℃放置过夜,然后在4 ℃下3 000 r/min离心10 min,取上清液用于测定肠道黏膜葡萄糖含量和二糖酶活性。
二糖酶活性(U/g)=Xn/30a。
式中:X为反应所释放的葡萄糖(μmol/L);a为二糖中葡萄糖释放量,麦芽糖为2,蔗糖和乳糖为1;n为样品稀释倍数;30为反应时间(min)。
1.3.3 肠道黏膜二糖酶mRNA表达丰度测定
1.3.3.1 总RNA的提取及引物设计
参照Trizol RNA抽提试剂盒(北京天根生物科技有限公司)说明书抽提肠道黏膜总RNA并采用核酸蛋白测定仪(GERMAN BiopHotometer 6131,Eppendorf公司,德国)测定总RNA的浓度和纯度,通过琼脂糖凝胶电泳检测总RNA的完整性。经过DNase-Ⅰ处理之后,按M-MLV试剂(Promega公司,美国)说明书进行反转录。反应结束后所获得的反应液即为合成的cDNA模板,-30 ℃保存备用。采用Primer 5.0软件设计鸡蔗糖酶-异麦芽糖酶(sucrase-isomaltase,SI)、麦芽糖酶-葡萄糖淀粉酶(maltase-glucoamylase,MGA)基因和内标基因β-肌动蛋白(β-actin)的上、下游引物。引物由上海生工生物工程有限公司合成,引物参数如表2所示。
1.3.3.2 二糖酶基因mRNA表达丰度的测定
采用实时荧光定量PCR方法测定二糖酶基因mRNA表达丰度。反应条件为:95 ℃ 1 min,(95 ℃ 15 s,58 ℃ 15 s,72 ℃ 15 s)×40个循环,(95 ℃ 1 min,58 ℃ 30 s,95 ℃ 30 s)×1个循环。反应体系见表3。目的基因mRNA表达丰度用2-ΔCt法进行计算。
ΔCt=(目的基因Ct-内标基因Ct);
mRNA表达丰度=2-ΔCt。
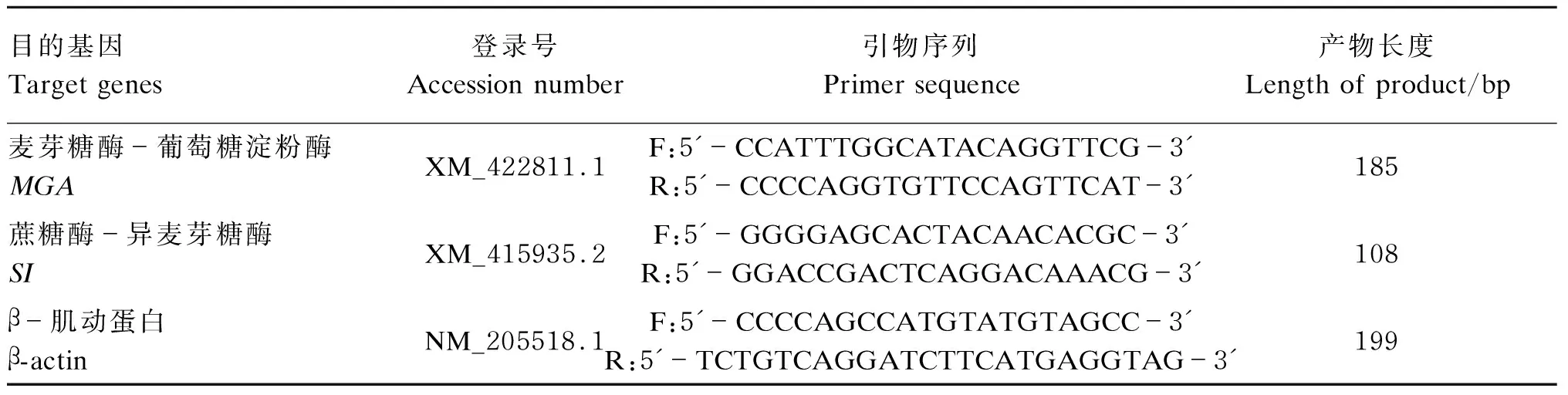
表2 引物参数

表3 实时荧光定量PCR反应体系
1.4 数据统计分析
数据采用SPSS 17.0软件进行统计分析,对肉鸡不同日龄不同肠段黏膜形态指标、黏膜生化指标以及二糖酶基因mRNA表达丰度分别进行单因素方差分析(one-way ANOVA),并采用LSD法进行多重比较。P<0.01视为差异极显著,P<0.05视为差异显著,结果均以平均值±标准误表示。
2 结果与分析
2.1 不同来源木聚糖酶及其组合对肉鸡肠道黏膜形态的影响
由表4可知,21日龄时,十二指肠绒毛宽度表现为Ⅰ、Ⅲ和Ⅳ组极显著大于Ⅱ组(P<0.01),Ⅳ组显著大于Ⅰ组(P<0.05);十二指肠隐窝深度表现为Ⅱ、Ⅲ和Ⅳ组极显著低于Ⅰ组(P<0.01);十二指肠V/C值表现为Ⅲ组极显著大于Ⅰ组(P<0.01),Ⅱ和Ⅳ组显著大于Ⅰ组(P<0.05)。21日龄时,空肠绒毛高度表现为Ⅳ组显著高于Ⅰ组(P<0.05);空肠隐窝深度表现为Ⅳ组显著低于Ⅰ组(P<0.05);空肠V/C值表现为Ⅱ和Ⅳ组极显著大于Ⅰ组(P<0.01),Ⅳ组显著大于Ⅲ组(P<0.05)。
42日龄时,十二指肠隐窝深度表现为Ⅱ和Ⅳ组极显著低于Ⅰ组(P<0.01),显著低于Ⅲ组(P<0.05);十二指肠V/C值表现为Ⅳ组极显著大于Ⅰ组(P<0.01),显著大于Ⅱ和Ⅲ组(P<0.05)。42日龄时,空肠绒毛高度表现为Ⅳ组极显著高于Ⅰ组(P<0.01),Ⅳ组显著高于Ⅱ组(P<0.05),Ⅲ组显著高于Ⅰ组(P<0.05);空肠隐窝深度表现为Ⅳ组显著低于Ⅰ和Ⅲ组(P<0.05);空肠V/C值表现为Ⅳ组极显著大于Ⅰ、Ⅱ和Ⅲ组(P<0.01),Ⅲ组显著大于Ⅰ组(P<0.05),Ⅲ组极显著大于Ⅱ组(P<0.01)。
63日龄时,肉鸡十二指肠和空肠绒毛高度、绒毛宽度和V/C值各组间差异均不显著(P>0.05)。十二指肠隐窝深度各组间差异不显著(P>0.05),而空肠隐窝深度则表现为Ⅱ、Ⅲ和Ⅳ组极显著低于Ⅰ组(P<0.01)。
2.2 不同来源木聚糖酶及其组合对肉鸡肠道黏膜生化指标的影响
2.2.1 不同来源木聚糖酶及其组合对肉鸡肠道黏膜葡萄糖含量的影响
由表5可知,21日龄时,肉鸡十二指肠中葡萄糖含量各组间差异不显著(P>0.05);空肠中葡萄糖含量Ⅲ和Ⅳ组均极显著高于Ⅰ和Ⅱ组(P<0.01),其中Ⅲ组最高,Ⅰ组最低。42日龄时,十二指肠中葡萄糖含量各组间差异不显著(P>0.05);空肠中葡萄糖含量Ⅳ组显著高于Ⅰ组(P<0.05)。63日龄时,十二指肠中葡萄糖含量各组间差异不显著(P>0.05);空肠中葡萄糖含量Ⅱ组极显著高于Ⅰ组(P<0.01),Ⅲ组显著高于Ⅰ组(P<0.05)。

表4 不同来源木聚糖酶及其组合对肉鸡肠道黏膜形态的影响
同行数据肩标无字母或相同字母表示差异不显著(P>0.05),相邻字母表示差异显著(P<0.05),相间字母表示差异极显著(P<0.01)。下表同。
In the same row, values with no letter or the same letter superscripts mean no significant difference (P>0.05), while with adjacent letter superscripts mean significant difference (P<0.05), and with alternate letter superscripts mean significant difference (P<0.01). The same as below.
2.2.2 不同来源木聚糖酶及其组合对肉鸡肠道黏膜麦芽糖酶活性的影响
由表6可知,21日龄时,麦芽糖酶活性在十二指肠中表现为Ⅳ组极显著高于Ⅰ、Ⅱ和Ⅲ组(P<0.01),Ⅲ组极显著高于Ⅰ组(P<0.01),Ⅱ组显著高于Ⅰ组(P<0.05);在空肠中表现为Ⅳ组极显著高于Ⅰ、Ⅱ和Ⅲ组(P<0.01),Ⅱ组极显著高于Ⅰ组(P<0.01)。42日龄时,麦芽糖酶活性在十二指肠中表现为Ⅱ和Ⅳ组极显著高于Ⅰ组(P<0.01),Ⅳ组显著高于Ⅲ组(P<0.05);在空肠中表现为各组间差异不显著(P>0.05)。63日龄时,麦芽糖酶活性在十二指肠中表现为Ⅲ和Ⅳ组极显著高于Ⅱ组(P<0.01),Ⅳ组显著高于Ⅰ组(P<0.05);在空肠中表现为各组间差异不显著(P>0.05)。

表5 21、42、63日龄十二指肠和空肠黏膜葡萄糖含量

表6 21、42、63日龄十二指肠和空肠黏膜麦芽糖酶活性
2.2.3 不同来源木聚糖酶及其组合对肉鸡肠道黏膜蔗糖酶活性的影响
由表7可知,21日龄时,蔗糖酶活性在十二指肠中表现为Ⅳ组极显著高于Ⅰ组(P<0.01),Ⅳ组显著高于Ⅲ组(P<0.05);在空肠中表现为Ⅳ组极显著高于Ⅰ、Ⅱ和Ⅲ组(P<0.01),Ⅲ组极显著高于Ⅰ组(P<0.01),Ⅱ组显著高于Ⅰ组(P<0.05)。42
日龄时,蔗糖酶活性在十二指肠中表现为Ⅳ组显著高于Ⅰ组(P<0.05);在空肠中表现为Ⅳ组极显著高于Ⅰ和Ⅱ组(P<0.01),Ⅳ组显著高于Ⅲ组(P<0.05),Ⅲ组显著高于Ⅰ组(P<0.05)。63日龄时,蔗糖酶活性在十二指肠和空肠均表现为各组间差异不显著(P>0.05)。

表7 21、42、63日龄十二指肠和空肠黏膜蔗糖酶活性
2.3 不同来源木聚糖酶及其组合对肉鸡肠道黏膜二糖酶基因mRNA表达丰度的影响
2.3.1 不同来源木聚糖酶及其组合对肉鸡肠道黏膜MGAmRNA表达丰度的影响
由表8可知,21日龄时,MGAmRNA表达丰度在十二指肠中表现为Ⅳ组显著高于Ⅰ组(P<0.05);在空肠中表现为Ⅱ组极显著高于Ⅰ组(P<0.01),Ⅲ和Ⅳ组显著高于Ⅰ组(P<0.05)。42日龄时,MGAmRNA表达丰度在十二指肠中表现为Ⅲ和Ⅳ组显著高于Ⅰ组(P<0.05);在空肠中表现为Ⅳ组极显著高于Ⅰ和Ⅱ组(P<0.01)。63日龄时,MGAmRNA表达丰度在十二指肠和空肠中均表现为各组间差异不显著(P>0.05)。

表8 不同来源木聚糖酶及其组合对肉鸡肠道黏膜MGA mRNA表达丰度的影响
2.3.2 不同来源木聚糖酶及其组合对肉鸡肠道黏膜SImRNA表达丰度的影响
由表9可知,21日龄时,SImRNA表达丰度在十二指肠中表现为Ⅳ组极显著高于Ⅰ组(P<0.01),Ⅱ组显著高于Ⅰ组(P<0.05);在空肠中表现为各组间差异不显著(P>0.05)。42日龄时,SImRNA表达丰度在十二指肠中表现为Ⅱ和Ⅳ组显著高于Ⅰ组(P<0.05);在空肠中表现为Ⅲ和Ⅳ组显著高于Ⅰ组(P<0.05)。63日龄时,SImRNA表达丰度在十二指肠中表现为Ⅳ组显著高于Ⅱ组(P<0.05);在空肠中表现为各组间差异不显著(P>0.05)。

表9 不同来源木聚糖酶及其组合对肉鸡肠道黏膜SI mRNA表达丰度的影响
3 讨 论
3.1 不同来源木聚糖酶及其组合对肉鸡肠道黏膜形态的影响
小肠绒毛形态结构的变化关系到小肠功能的发挥,也决定了其对营养物质的消化吸收能力。肠道绒毛上皮细胞主要发挥吸收功能,隐窝上皮细胞主要发挥分泌功能,肠道净吸收取决于肠道黏膜V/C值的高低。肠道黏膜V/C值上升,则黏膜功能改善,消化吸收功能增强,生长发育加快[9]。
小麦饲粮中的非淀粉多糖能够影响动物肠道黏膜形态[10]。研究表明,木聚糖酶的添加能够不同程度的影响动物肠道黏膜绒毛高度、隐窝深度以及V/C值[11-13]。而Yang等[14]报道,小麦饲粮添加木聚糖酶并没有显著提高肉仔鸡空肠绒毛高度,但是降低了空肠隐窝深度。本试验中,在小麦-豆粕型饲粮中添加木聚糖酶能够不同程度地增加小鸡和中鸡十二指肠和空肠绒毛高度和V/C值,且不同程度地降低小鸡(21日龄)和中鸡(42日龄)十二指肠和空肠黏膜隐窝深度,与前人报道基本一致。
在本试验中,木聚糖酶对小鸡(21日龄)、中鸡(42日龄)和大鸡(63日龄)的不同肠段绒毛高度、绒毛宽度和隐窝深度的影响效果不同,其原因可能与动物所处的生理阶段不同以及酶制剂本身的因素有关。
3.2 不同来源木聚糖酶及其组合对肉鸡肠道黏膜二糖酶活性的影响
上皮细胞不断从绒毛顶端衰亡脱落的过程,实质上是二糖酶及其他低聚糖酶的分泌释放过程。淀粉进入小肠后,首先经过胰淀粉酶作用产生α-糊精、麦芽丙糖和麦芽糖等二糖,随后在刷状缘表面二糖酶等作用下产生直接被肠绒毛吸收的单糖。因此,肠道黏膜葡萄糖含量能够在一定程度上反映碳水化合物在黏膜上的消化能力,也能间接反映出二糖酶的性。木聚糖酶可能通过影响食糜黏性达到减小相对不动水层厚度,从而影响肠道黏膜二糖酶活性和葡萄糖含量。
麦类饲粮中含有可溶性非淀粉多糖,能够影响动物肠道黏膜二糖酶活性[15]。非哺乳动物肠道黏膜中的二糖酶主要以麦芽糖酶和蔗糖酶为主。许梓荣等[16]报道,饲喂添加非淀粉多糖酶饲粮的仔猪空肠黏膜麦芽糖酶和蔗糖酶活性较饲喂不添加非淀粉多糖酶饲粮的仔猪分别提高了38.46%和40.00%。本试验中,木聚糖酶在小鸡和中鸡阶段很大程度上提高了肠道黏膜中蔗糖酶和麦芽糖酶的活性,与上述报道结果一致。木聚糖酶对肠道黏膜二糖酶活性产生影响的机制可能是:1)降解小麦饲粮中阿拉伯木聚糖,为二糖酶提供更多的作用底物;2)降低水溶性非淀粉多糖黏性,从而降低小肠食糜的黏度,使得麦芽糖、蔗糖等二糖更容易到达水解位点,促进二糖的水解。
3.3 不同来源木聚糖酶及其组合对肉鸡肠道黏膜二糖酶基因表达的影响
动物肠道黏膜中二糖酶基因mRNA表达丰度随着动物发育而变化。翁梅倩等[17]报道,新生兔乳糖根皮苷水解酶(lactase-phlorizin hydrolase,LPH)和SImRNA在各肠段都有表达,完全断乳15 d后其表达明显增强。但Yadgary等[18]有不同的报道,胚胎中11~21日龄的小鸡SImRNA表达丰度未随着日龄的增长而显著增加。本试验测定了肉鸡3个生长阶段肠道黏膜中MGA和SImRNA的表达丰度,但未发现统一的规律。动物饲粮组成、激素等均能影响肠道黏膜二糖酶基因的表达。Mochizuki等[19]报道,小鼠饲喂高淀粉低脂肪饲粮后空肠麦芽糖酶和葡萄糖淀粉酶的活性升高。本试验在小麦-豆粕型饲粮中添加木聚糖酶,在小鸡和中鸡阶段不同程度地增强了肠道黏膜中MGA和SImRNA的表达丰度。
3.4 组合酶设计的理论基础及其意义
组合酶是指利用酶催化的协同作用,在催化水解同一底物的不同来源和特性的酶中选择具有互补性的2种或2种以上酶配合而成的酶制剂[7]。本试验挑选7种不同来源的单一木聚糖酶,分别测定其酶学特性,再根据性质的不同,挑选出具有互补性的单一木聚糖酶1#和单一木聚糖酶2#,进行不同比例的组合。然后在模拟肉鸡胃肠道环境的情况下,根据木聚糖酶的酶活特性进行筛选,选出具有互补效果最佳的模型。随着非常规饲料原料的大量使用,高效的组合酶将更有优势[20]。因此,今后饲料酶制剂的发展方向,应该是研发和使用更多的组合酶或组合型复合酶产品。
4 结 论
肉鸡在前(1~21日龄)、中期(22~42日龄)阶段时,木聚糖酶的添加能够改善肠道黏膜形态,促进二糖酶的分泌及其基因的表达;在活性相同的情况下,2个单一木聚糖酶的作用效果相近,而组合酶的作用效果则优于单酶,表现出高效催化的组合效应。
致谢:
衷心感谢广东温氏食品集团试验鸡场在动物饲养试验方面提供的便利和帮助。
[1] FRIESEN O D,GUENTER W,MARQUARDT R R,et al.The effect of enzyme supplementation on the apparent metabolizable energy and nutrient digestibilities of wheat,barley,oats,and rye for the young broiler chick[J].Poultry Science,1992,71(10):1710-1721.
[2] BRENES A,MARQUARDT R R,GUENTER W,et al.Effect of enzyme supplementation on the nutritional value of raw,autoclaved,and dehulled lupins (Lupinusalbus) in chicken diets[J].Poultry Science,1993,72(12):2281-2293.
[3] SIEO C C,ABDULLAH N,TAN W S,et al.Influence of β-glucanase-producingLactobacillusstrains on intestinal characteristics and feed passage rate of broiler chickens[J].Poultry Science,2005,84(5):734-741.
[4] UNI Z,NOY Y,SKLAN D.Posthatch development of small intestinal function in the poultry[J].Poultry Science,1999,78(2):215-222.
[5] WU Y B,RAVINDRAN V,THOMAS D G,et al.Influence of method of whole wheat inclusion and xylanase supplementation on the performance,apparent metabolisable energy,digestive tract measurements and gut morphology of broilers[J].British Poultry Science,2004,45(3):385-394.
[6] SENKOYLU N,SAMLI H E,AKYUREK H,et al.Effects of whole wheat with or without xylanase supplementation on performance of layers and digestive organ development[J].Italian Journal of Animal Science,2009,8(2):155-163.
[7] 冯定远.饲料工业的技术创新与技术经济[J].饲料工业,2004(11):1-6.
[8] FRANKEL W L,ZHANG W,AFONSO J,et al.Glutamine enhancement of structure and function in transplanted small intestine in the rat[J].Journal of Parenteral and Enteral Nutrition,1993,17(1):47-55.
[9] 王子旭,佘锐萍,陈越,等.日粮锌硒水平对肉鸡小肠黏膜结构的影响[J].中国兽医科技,2003,33(7):18-21.
[10] 贺永惠,王清华,王艳荣,等.小麦非淀粉多糖与木聚酶对大鼠小肠形态结构的影响[J].河南科技学院学报(自然科学版),2010,38(2):56-59.
[11] 范成莉.β-葡聚糖、木聚糖复合酶对断奶仔猪生长轴激素的影响及其机理研究[D].博士学位论文.杭州:浙江大学,2008
[12] 丁雪梅,张克英.玉米-杂粕型日粮添加木聚糖酶对肉鸡免疫功能、肠道形态和微生物菌群的影响[J].中国畜牧杂志,2010,46(23):26-30.
[13] GHAYOUR-NAJAFABADI P,KHOSRAVINIA H,GHEISARI A,et al.Productive performance,nutrient digestibility and intestinal morphometry in broiler chickens fed corn or wheat-based diets supplemented with bacterial- or fungal-originated xylanase[J].Italian Journal of Animal Science,2017:doi:10.1080/1828051X.2017.1328990.
[14] YANG Y,IJI P,KOCHER A,et al.Effects of xylanase on growth and gut development of broiler chickens given a wheat-based diet[J].Asian-Australasian Journal of Animal Sciences,2008,21(11):1659-1664.
[15] 孙哲,汪儆,雷祖玉,等.小麦可溶性非淀粉多糖对肉仔鸡小肠粘膜二糖酶活性的影响[J].动物营养学报,2003,15(1):26-30.
[16] 许梓荣,李卫芬,孙建义.NSP酶对大麦体外消化的影响[J].中国粮油学报,2002,17(2):51-54.
[17] 翁梅倩,夏红萍,钱龙华,等.兔肠发育中与糖消化吸收相关基因的表达[J].上海第二医科大学学报,2001,21(6):495-499.
[18] YADGARY L,YAIR R,UNI Z.The chick embryo yolk sac membrane expresses nutrient transporter and digestive enzyme genes[J].Poultry Science,2011,90(2):410-416.
[19] MOCHIZUKI K,HONMA K,SHIMADA M,et al.The regulation of jejunal induction of the maltase-glucoamylase gene by a high-starch/low-fat diet in mice[J].Molecular Nutrition & Food Research,2010,54(10):1445-1451.
[20] 冯定远.新型高效饲料组合酶的最新理论研究与应用技术[J].饲料工业,2011,32(4):1-8.
