模拟氮沉降对云杉人工林土壤有机碳组分及理化性质的影响
2018-06-07乔枫史伟薛元杰
乔枫,史伟,薛元杰
1. 国家林业局西北林业调查规划设计院,陕西 西安 710048;2. 中国科学院成都生物研究所生态恢复重点实验室,四川 成都 610041
碳循环在地球生态系统的物质循环与能量交换中起着重要作用。土壤作为植被生长的主要载体,在碳循环系统中扮演着重要的角色(王绍强等,1999)。土壤有机碳是植被碳库的2.5倍,制约着植物的光合作用和生长发育,是生物化学循环中重要的生态因子(王绍强等,2000;彭新华等,2004)。土壤有机碳的组成形态不同,其土壤的肥力影响也不尽一致;土壤有机碳能够直接影响土壤微生物新陈代谢等活动,进而影响植物的生长。另外,土壤微生物也能够产生生物碳,虽然数量较少,但能够参与整个生态循环,进而促进土壤养分循环,加速腐殖质分解。不同土壤有机碳组分在土壤循环过程中发挥着各自的作用,因此具体探究不同有机碳组分,能够更好地研究土壤有机碳质量,从而为改善土壤结构和养分等提供有益参考(张英英等,2017)。活性有机碳是土壤有机碳中最为活跃的那一部分碳,能够充分被微生物吸收利用,同时能够改善土壤结构、提升植物养分供给,对促进植物的生长具有显著的作用,因此,研究土壤活性有机碳有助于更好地研究土壤结构、增强植物养分供给,反映有机碳的抗氧化程度和利用难易程度(张忠启等,2015;廖丹等,2015)。
随着工业化的迅猛发展和人类活动的加剧,各类含氮化合物(氮肥、汽车尾气等)被大量排放,尤其是煤等矿物质燃烧产生了大量碳氮化合物,大气氮沉降问题日益突出,已成为全球性问题(刘彩霞等,2015)。中国已然成为全球第三大氮沉降危险区(李德军等,2015;王晶苑等,2013)。就全球而言,氮沉降已然成为整个生态系统面临的生态现象,氮沉降量的增加为森林的生态循环带来了新的挑战(张炜等,2015)。长期的高氮输入显著提升了土壤的矿化速度,加剧土壤的酸化,进而带来土壤板结、养分不足等问题,不利于植物的生长发育(樊后保等,2007;段洪浪等,2009;陈浩等,2012;方华等,2015)。当前,国内外有关氮沉降对森林生态系统碳循环影响的研究已经引起了足够大的关注,这对于碳循环的研究具有重要的意义(陈美领等,2016)。例如,Magnani et al.(2007)认为南方和北方的土壤具有较为显著差异的碳含量,但这种差异能够较好地通过氮沉降进行相应的解释。国内学者樊后保等(2015)通过对福建国有林场进行长期的观测研究分析发现,氮沉降对杉木Cunninghamia lanceolata林生态循环具有重要的影响作用。莫江明等(2004)将鼎湖山的马尾松Pinus massoniana及混交林作为研究对象,探究氮沉降在森林生态循环中的主要影响效果,并对其影响机理进行了分析。然而,在众多的研究中,对于深入到氮沉降对森林生态系统土壤碳组分的研究尚不多见,而碳组分,特别是土壤活性有机碳,只要发生微小的变化,就可对森林生态系统的碳循环产生重要作用,因此,深入探究氮沉降对森林生态系统土壤碳组分的研究对于研究生态系统碳循环具有重要意义。
大气氮素以无机态和有机态形式发生沉降,二者分别占总氮沉降量的70%和30%左右(方运霆等,2004)。其中,无机氮素形态主要以硝态氮和铵态氮为主,而有机氮沉降形态主要为尿素和氨基酸(李德军等,2003)。有研究表明,与无机氮相比,施用有机氮其生态系统环境受到的干扰相对较小(李德军等,2003)。此外,与矿化、硝化作用一样,氮施加引起的反硝化作用的影响与施加氮的类型有关,尿素比其他氮肥更容易刺激反硝化作用的发生,可能是因为:(1)尿素促进硝化作用而提供了硝酸盐源;(2)尿素提高了土壤pH值;(3)尿素水解使含碳化合物向可溶性方向转换,进而为反硝化菌提供能量源。另一方面,矿质氮肥施加和随后的盐度增加可能引起渗透胁迫而抑制异养微生物的活动(王晖等,2008;吴迪等,2015)。
四川的高山针叶林作为长江重要的生态屏障之一,在上世纪中叶,遭受了大量采伐,虽然后续也种植了大量的人工林,但因其过度采伐,超过了生态承受能力,导致大面积自然林消失。其中人工林以大面积的粗枝云杉Picea asperata为主,形成了较为单一的树种结构,物种多样性水平低下,其土壤肥力远不如原始林(庞学勇等,2003)。目前多从地上部分对川西地区针叶林开展相应的研究,对土壤的研究也多从养分方面开展,而有关氮沉降对土壤有机碳组分的影响尚不多见。结合华南地区前人关于氮沉降的研究,本研究采用尿素进行氮沉降模拟,研究氮沉降对川西针叶林土壤有机碳组分的影响,以期为该区土壤优化和林业资源管理提供参考。
1 材料与方法
1.1 研究区概况
本实验区位于四川阿坝地区的米亚罗镇(31°11'~31°47'N,102°35'~103°31'E),属于岷江上游,因其地形地貌和气候特殊,形成了较为独特的山地季风气候,四季分明。冬天虽然较为寒冷,但日照时间较充足;全年降雨集中在夏季,年均降雨量近800 mm。该地区海拔高达3300 m,坡度约为23°,平均气温约为 9 ℃。该地区的原生树种主要是岷江云杉 Picea asperata,属于亚高山针叶林,生长在2400 m以上地区,上世纪50年代以后原始森林被大量采伐,故而形成了悬钩子 Rubus crataegifolius及箭竹Pseudosasa japonica等灌丛,后进行了粗枝云杉的人工种植。本研究于2015年7月开展了相应的底值调查,具体结果如表1所示。
模拟氮沉降实验即以人工喷氮的方式对未来的氮沉降趋势进行模拟。本研究参照前人类似研究设计(Fenn et al.,1998),按氮施用量的高低,设置4种处理,采用随机区组设计,从低到高分别标记为 N0(0 kg·hm-2·a-1,对照),N1(60 kg·hm-2·a-1),N2(120 kg·hm-2·a-1),N3(240 kg·hm-2·a-1),每种处理重复3次(样地面积为100 m×100 m),样地之间留10 m宽的缓冲带。2015年12月建立样地后,于2016年1月开始进行模拟氮沉降处理,每月月初以溶液的形式将氮素喷洒于样地地面。按照处理水平的要求,将尿素[CO(NH2)2]溶解在20 L水中,以背式喷雾器在林地人工来回均匀喷洒。对照样地喷施等量水,以减少因处理之间的差异。
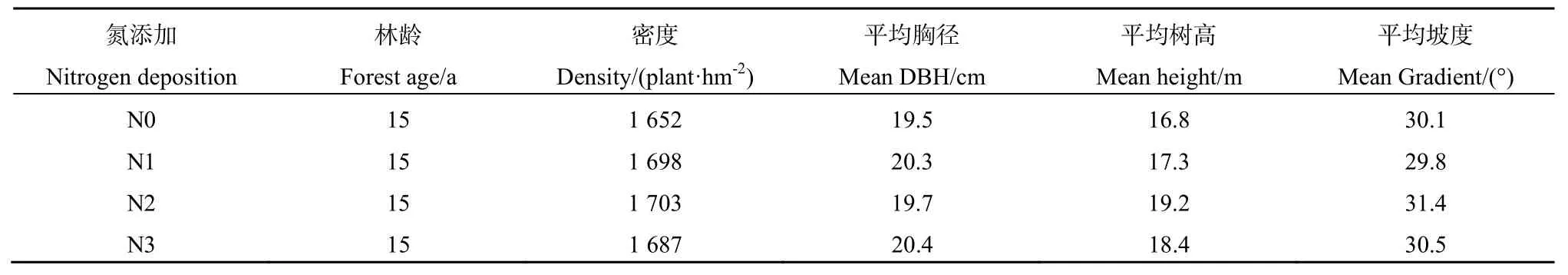
表1 各样地的主要林分特征Table 1 The characteristics of the sampling sites
1.2 土壤样品采集
于2017年9月中旬,对4种样地进行取样,每个样地内随机设置5个采样点,每个采样点间距在10 m以上,每个采样点重复取5个土壤样品作为平行,5个平行样品之间间隔2 m,为了保证取样的一致性,所取样的土壤坡度均小于5°。采用四分法留取1 kg左右土样,取样深度为0~20 cm混合土样(除去表层的枯落物层);所取样品分为两部分,一部分鲜土现场过2 mm筛后于4 ℃保温冰箱中保存用于测定土壤微生物量,另一部分带回实验室风干后去除杂质后对其养分和有效养分进行测定。在取样点附近挖取剖面测定土壤容重(环刀法,g·cm-3),并计算土壤总孔隙度(%)。
1.3 土壤样品测定
土壤理化性质及养分含量的测定:土壤容重(BD)采用烘干法,计算土壤孔隙度(TPO);机械组成采用激光粒度分析仪测定;pH采用电极电位法(1∶2.5土水比)测定;土壤有机碳(SOC)采用重铬酸钾氧化外加热法测定;土壤全氮(TN)采用全自动凯氏定氮法测定;土壤全磷(TP)采用NaOH熔融-钼锑抗比色法测定;有效磷(AP)采用 NaHCO3浸提-钼锑抗比色法测定;全钾(TK)和有效钾(AK)采用火焰分光光度法测定(鲍士旦,2000)。
土壤有机碳组分的分离:将土壤研磨过 0.25 mm筛,60 ℃水浴锅蒸干,然后60 ℃烘干12 h,冷却,称重。易氧化有机碳(EOC)采用 K2MnO4氧化-比色法测定(张俊华等,2010);颗粒有机碳(POC)采用湿筛法测定(黄雪夏等,2007);轻质有机碳(LFOC)采用浮选法测定(杨君珑等,2017);水溶性有机碳(WSOC)采用TOC分析仪测定(Phoenix 8000,Teledyne Tekmar公司,美国)(曾婷婷等,2016);土壤微生物量碳采用氯仿熏蒸提取法测定(SMBC),熏蒸和未熏蒸处理的碳含量之差,再乘以转换系数K(EC)即可得到生物量碳,即BC=2.64EC(吴金水,2006)。
1.4 统计分析
利用Excel 2003和SPSS 18.00软件对数据进行单因素方差分析(One-way ANOVA),采用邓肯新复检验法进行差异显著性检验。所有数据测定结果以平均值±标准误的形式表达。通径分析(Path Analysis)用于研究自变量(土壤理化性质和养分)和因变量(土壤有机碳组分)之间的相互关系、作用方式。通过通径分析,能够找出自变量对因变量影响的直接效应和间接效应。为了进一步研究在氮添加模式下土壤理化性质和养分对土壤有机碳组分的影响,于SPSS软件中对土壤理化性质和养分进行通径分析,明确其对有机碳组分的贡献程度。
2 结果与分析
2.1 氮沉降对土壤粒径组成及理化性质的影响
表2反映了氮沉降对人工林土壤机械组成及理化性质的影响,其中1~0.05 mm土壤所占比例范围为18.25%~21.79%,平均值为19.89%,基本表现为:N3>N2>N1>N0,其中N3>N2差异不显著(P>0.05),显著高于N0(P<0.05);0.05~0.002 mm土壤所占比例范围为 54.60%~56.86%,基本表现为:N3>N2>N1>N0,其中 N3和 N2差异不显著(P>0.05),二者显著高于 N1和 N0(P<0.05);<0.002 mm土壤所占比例范围为21.35%~26.98%,基本表现为:N3>N2>N1>N0,其中N0和N1差异不显著(P>0.05),二者显著高于 N2 和 N3(P<0.05)。BD变化范围在1.03~1.26 g·cm-3之间,随N处理水平的增加而降低,其中N0和N1显著高于N2和N3(P<0.05),而N2和N3差异不显著(P>0.05);TPO与BD变化趋势相反,随N处理水平的增加而增加,其中N0、N1和N2差异均不显著,三者显著低于 N3(P<0.05);土壤 pH值变化范围在6.58~7.02之间,随 N处理水平的增加而降低,其中N1和N2显著高于N3和N4(P<0.05)。
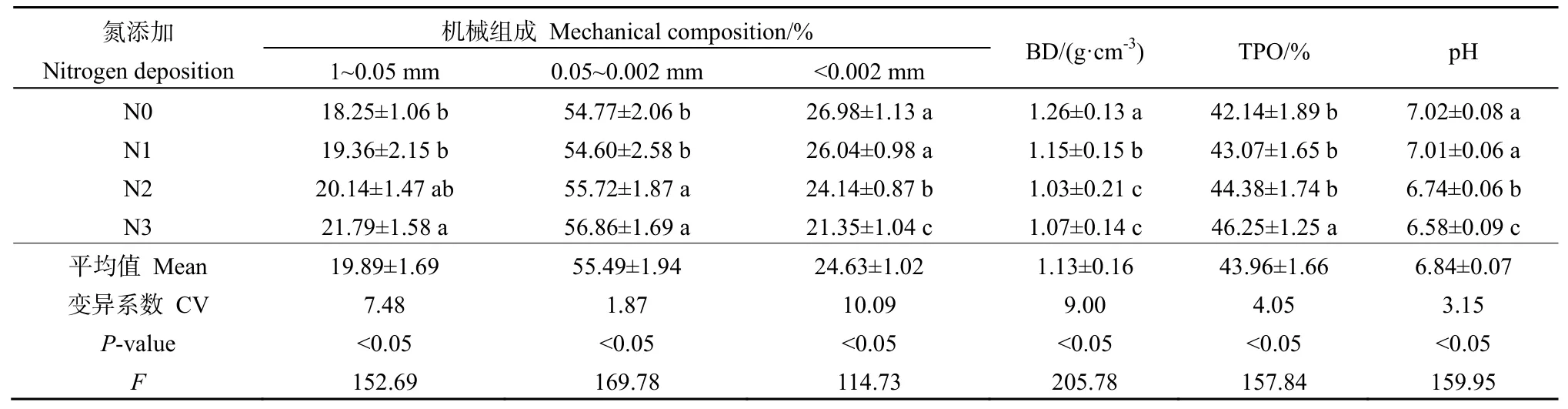
表2 氮沉降对土壤粒径组成及理化性质的影响Table 2 Effects of simulated nitrogen deposition on soil mechanical composition and physico-chemical properties in Picea asperata plantation in Sichuan
2.2 氮沉降对土壤养分含量的影响
由表3可知,在N添加基础上,土壤养分(除了 TP)和有效养分均呈现出一致性规律,随着 N处理水平的增加而增加。土壤有机碳质量分数变化范围为 12.56~16.78 g·kg-1,平均值为 15.51 g·kg-1,其中N2和N3差异不显著(P>0.05),N0和N1差异不显著(P>0.05),N2和N3显著高于N0和N1(P<0.05);TN 变化范围为 0.98~1.67 g·kg-1,平均值为 1.29 g·kg-1,不同 N 处理下差异均显著(P<0.05);TK 变化范围为 15.23~29.78 mg·kg-1,平均值为21.26 mg·kg-1,其中N1和N0差异不显著(P>0.05),二者显著低于 N2和 N3(P<0.05);AP变化范围为20.08~39.78 mg·kg-1,平均值为26.94 mg·kg-1,不同 N 处理下差异均显著(P<0.05);AK变化范围为75.23~91.07 mg·kg-1,平均值为8.22 mg·kg-1,其中 N1和 N0差异不显著(P>0.05),N3显著高于N1和N0(P<0.05)。
2.3 氮沉降对土壤有机碳组分的影响
氮沉降对土壤活性、轻组和颗粒有机碳含量的影响见表4,由表可知,EOC、POC、LFOC、WSOC和SMBC明显受氮沉降的影响。EOC、POC、LFOC、WSOC均呈现出一致性规律,随N处理水平的增加而增加。EOC变化范围为1.26~1.69 mg·kg-1,平均值为 1.46 g·kg-1,其中 N0和 N1差异不显著(P>0.05),二者显著低于 N2和 N3(P<0.05);POC变化范围为1.42~1.72 mg·kg-1,平均值为1.54 mg·kg-1,其中 N0和 N1差异不显著(P>0.05),二者显著低于N2和N3(P<0.05);LFOC变化范围为 1.32~2.23 mg·kg-1,平均值为 1.80 mg·kg-1,不同处理下差异均显著(P<0.05);WSOC变化范围为 6.98~12.56 mg·kg-1,平均值为 9.37 mg·kg-1,其中N0和N1差异不显著(P>0.05),二者显著低于N2和N3(P<0.05);SMBC变化范围为69.85~197.46 mg·kg-1,平均值为 120.70 mg·kg-1,不同处理下差异均显著(P<0.05)。
2.4 土壤活性碳与总有机碳的比例关系
易氧化有机碳分配比例是指其占土壤有机碳含量的多少,可用于表征土壤有机碳的稳定性,比例越高,说明土壤碳稳定性越差。由表 5可知,EOC/SOC比例变化范围为10.03%~13.46%,变异系数范围在8.56%~12.47%之间,基本表现为N3>N2>N1>N0,其中N1和N0差异不显著(P>0.05),二者显著低于N3和N2(P<0.05),说明氮沉降对土壤EOC/SOC有显著影响。土壤微生物量碳占有机碳的百分比称为微生物熵。微生物熵的变化反映了土壤中输入的有机质向微生物量碳的转化效率、土壤中碳损失和土壤矿物对有机质的固定。土壤SMBC/SOC比例变化范围在0.56%~1.57%,基本表现为 N3>N2>N1>N0,其中不同处理间的 SMBC/SOC比例差异均显著(P<0.05)。
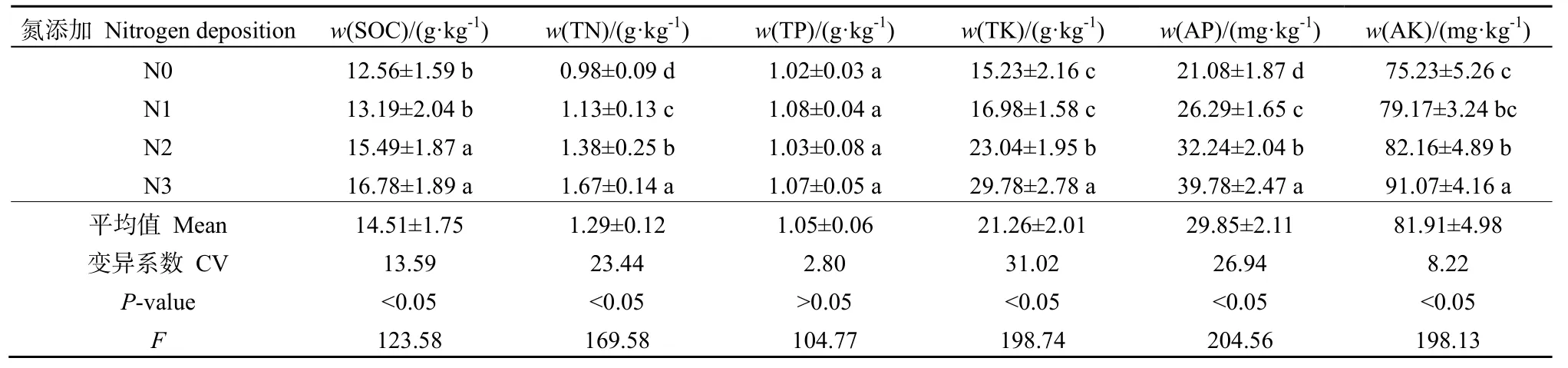
表3 氮沉降对土壤养分含量的影响Table 3 Effects of simulated nitrogen deposition on soil nutrients in Picea asperata plantation in Sichuan
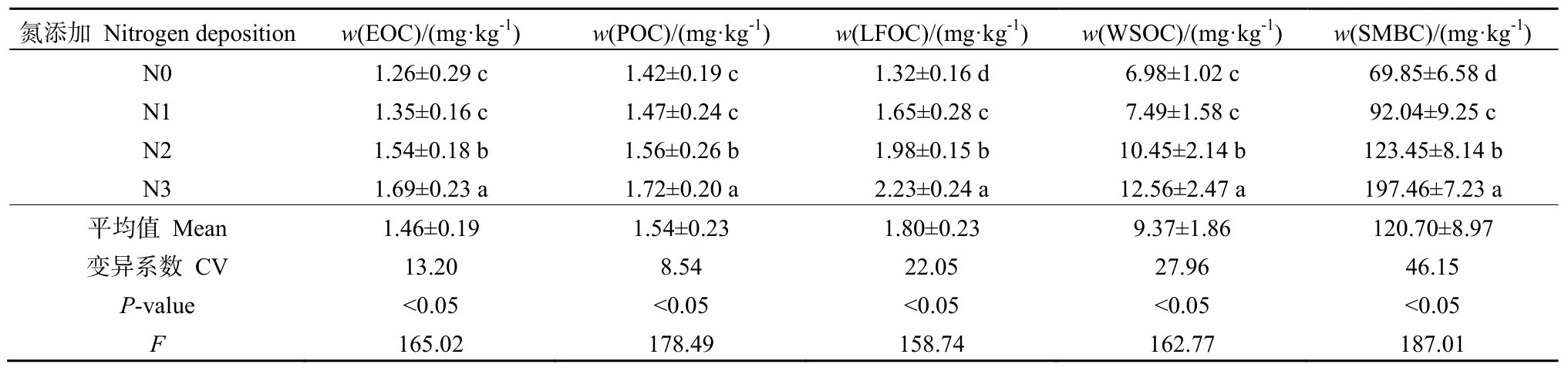
表4 氮沉降对土壤碳组分的影响Table 4 Effects of simulated nitrogen deposition on soil organic carbon fractions in Picea asperata plantation in Sichuan
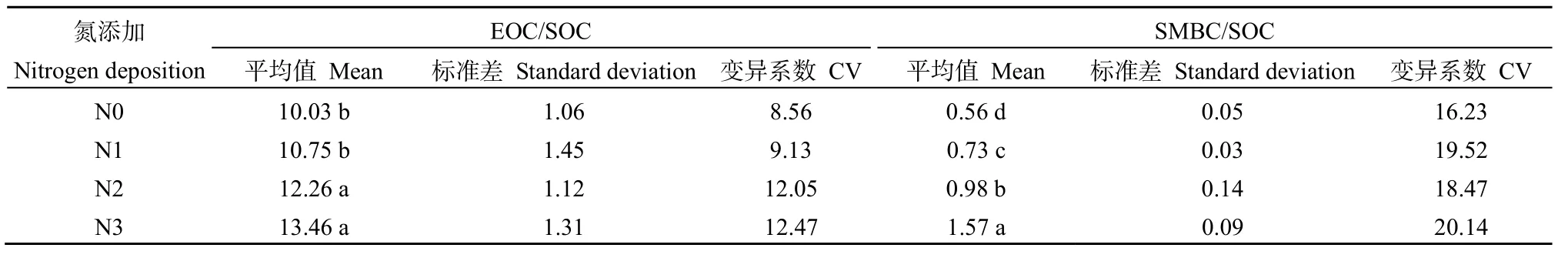
表5 EOC/SOC和SMBC/SOC比例Table 5 Effects of simulated nitrogen deposition on the ratio of EOC/SOC and SMBC/SOC
2.5 土壤有机碳组分与土壤环境因子的通径分析
表6和表7反映了环境因子对土壤EOC、POC、LFOC、WSOC和SMBC影响的通径系数。由表6可知,1~0.05 mm粒径和TPO对土壤有机碳组分产生直接效应;<0.002 mm和pH对土壤有机碳组分产生间接效应;土壤理化性质对土壤有机碳组分产生的总效应值具体表现为 1~0.05 mm>pH>TPO=(<0.002) mm>BD>0.05~0.002 mm。
表7反映了土壤养分对土壤有机碳组分影响的通径系数。不同养分对土壤有机碳组分产生直接和间接负作用,SOC、TN和 AK对土壤有机碳组分产生直接效应;TK和AP对土壤有机碳组分产生间接效应;总效应值大小依次为 SOC>TN>AK>TK>TP>AP。
3 讨论
本研究中氮沉降对土壤理化性质和养分的影响并不一致,土壤孔隙度与其容重具有相反变化趋势,土壤pH随着N浓度的增加而降低,与此同时,氮沉降增加了土壤养分含量,对TP没有明显的影响,氮沉降通过降低土壤pH值,增大土壤孔隙度,促进了土壤微生物代谢活动,使养分增加,进而促进了土壤有机碳的分解(邬建红等,2015;陈志杰等,2016)。Schuur et al.(2015)研究发现,土壤微生物在土壤有机碳的分解中起着重要的作用,尤其是微生物的数量及活性。本研究中,SOC、TN和pH对POC的直接作用最大,pH值的降低导致有机碳各组分的增加。由此可知,在氮沉降处理下,pH值的降低和孔隙度的增加,加速了微生物的代谢活动,氮沉降在一定程度上促进了SOC的增加,有利于土壤有机碳的累积。
有研究发现,高浓度N并没有抑制土壤微生物量碳,在正常水平下,土壤微生物量碳呈现增长的变化趋势,当浓度较低氮进入土壤后微生物能够及时将之消化吸收,但是短时间内如果高浓度氮进入土壤,则很容易导致微生物数量下降,多余的氮难以被及时消化吸收,很可能以 NO3-的方式形成淋溶,最终导致土壤酸化,酸化使Al3+等有毒离子释放从而形成铝毒,制约微生物的生长及活动(许凯等,2014)。长时间的氮输入很容易导致微生物量碳减少,且氮影响生物量分配,制约着土壤的理化特性,土壤生物量下降和土壤酸化问题也将更加明显,对地下微生物新陈代谢等活动形成显著抑制作用。一定量的碳氮进入土壤将为微生物提供更为充足的可吸收碳氮,从而有利于微生物的新陈代谢等活动(林伟等,2016)。龙凤玲等(2014)研究发现,对于多数人工林而言,一般处于氮限制的状态,当氮限制缓解时,植物会为地上部分分配更多的碳,地下微生物量会发生相应的下降。就人工林生态系统而言,微生物群落具有相对稳定的结构,能够更好地应对氮素沉降,在氮沉降过程中能够发生较为明显的固氮作用。
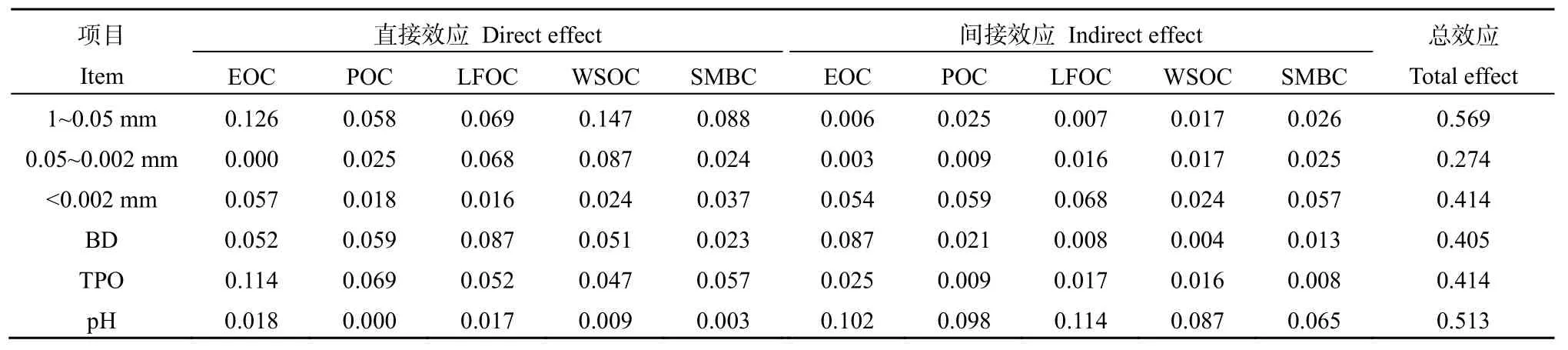
表6 土壤有机碳组分与土壤理化性质的通径分析Table 6 Pathway analysis between soil organic carbon fractions and soil physico-chemical properties
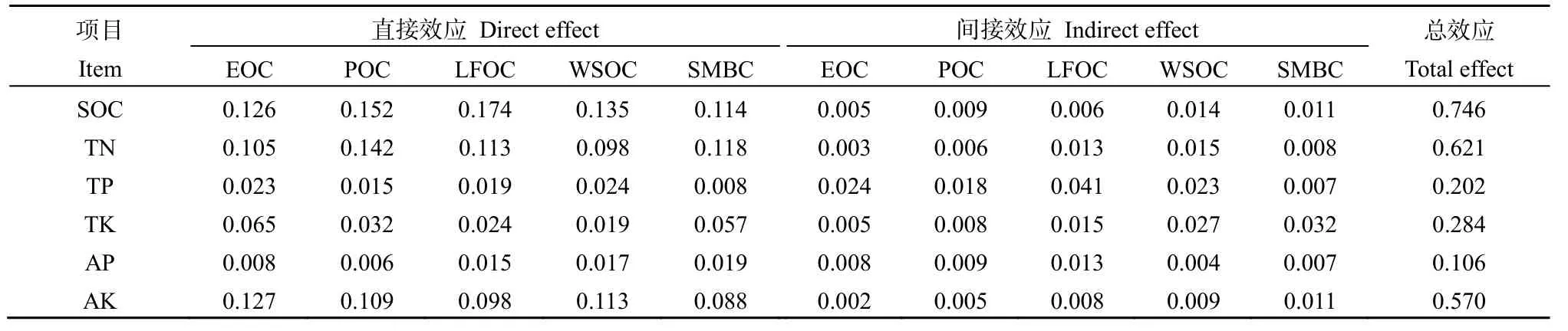
表7 土壤有机碳组分与土壤养分的通径分析Table 7 Pathway analysis between soil organic carbon fractions and soil nutrients
大量研究发现,枯枝落叶及生物残体等一定量的腐殖质分解能够显著促进土壤有机碳的积累,提高土壤肥力(张建华等,2017)。本研究中,N输入增加了枯枝落叶等凋落物,为微生物提供了更多的能量,从而释放更多的可溶性有机碳,另外,微生物数量的增多也能够促进腐殖质的分解速度和质量,提升土壤肥力,改善土壤结构;此外,根的活动加速,根系分泌速度显著加快,能够产生更多的有机碳(游成铭等,2016)。例如,全权等(2015)研究发现,氮沉降能够对土壤木质素的分解形成抑制作用,进而提升土壤有机碳含量。马芬等(2015)认为,氮沉降能够对可溶性酚类物质积累起促进作用,也能够促进可溶性碳的增加。经过长时间的氮输入发现,有机碳的淋溶会相应加快,含量降低。通径分析结果表明,土壤有机碳组分的影响因素具有多样性,其中1~0.05 mm粒径和TPO能够对之产生直接的作用,而0.002 mm以下粒径及pH间接影响有机碳组分;土壤养分对有机碳组分产生的是负向影响,且SOC、TN、AK能够产生直接负作用,TK、AP则产生间接的负向影响。因此,氮沉降能够促进云杉人工林有机碳组分的稳定性,对有机碳积累具有促进作用。
4 结论
(1)氮沉降对土壤理化性质和养分的影响较为明显,随着氮浓度的增加,土壤总孔隙度(TPO)和土壤容重(BD)呈相反的变化趋势,土壤养分(有机碳SOC、全氮TN、全钾TK、有效磷AP和有效钾AK)随着N浓度的增加而增加,说明氮沉降对土壤养分起到了一定的促进作用。然而,长期的氮沉降是否仍然对土壤养分起促进作用,还需要后续试验进行验证。
(2)土壤易氧化有机碳(EOC)、颗粒有机碳(POC)、轻组有机碳(LFOC)、水溶性有机碳(WSOC)、微生物量碳(SMBC)、EOC/SOC比例和微生物熵(MBC/SOC)均随N浓度的增加而增加。通径分析表明,SOC、TN和 AK对土壤有机碳组分产生直接效应,TK和AP对土壤有机碳组分产生间接效应,说明氮沉降能够促进云杉人工林有机碳组分的稳定性,有效养分对有机碳积累贡献更大,其机理还有待进一步研究。
FENN M E, POTH M A, ABER J D, et al. 1998. Nitrogen excess in north american ecosystems: predisposing factors, ecosystem responses, and management strategies. Ecological Applications, 8(3): 706-733.
HYVONEN R, PERSSON T, ANDERSSON S, et al. 2008. Impact of long-term nitrogen addition on carbon stocks in trees and soils in northern Europe [J]. Biogeochemistry, 89(1): 121-137.
MAGNANI F, MENCUCCINI M, BORGHETTI M, et al. 2007. The human footprint in the carbon cycle of temperate and boreal forests [J].Nature, 447(7146): 849-851.
SCHUUR E A G, MCGUIRE A D, SCHADEL C, et al. 2015. Climate change and the permafrost carbon feedback [J]. Nature, 520(7546):171-179.
鲍士旦. 2000. 土壤农化分析[M]. 北京: 中国农业出版社.
曾婷婷, 张玲玲, 李意德, 等. 2016. 林型转化对土壤pH、有机碳组分和交换性矿质元素的影响[J]. 生态环境学报, 25(4): 576-582.
陈浩, 莫江明, 张炜, 等. 2012. 氮沉降对森林生态系统碳吸存的影响[J]. 生态学报, 32(21): 6864-6879.
陈美领, 陈浩, 毛庆功, 等. 2016. 氮沉降对森林土壤磷循环的影响[J].生态学报, 36(16): 4965-4976.
陈志杰, 韩士杰, 张军辉. 2016. 土地利用变化对漳江口红树林土壤有机碳组分的影响[J]. 生态学杂志, 35(9): 2379-2385.
段洪浪, 刘菊秀, 邓琦,等. 2009. CO2浓度升高与氮沉降对南亚热带森林生态系统植物生物量积累及分配格局的影响[J]. 植物生态学报,33(3): 570-579.
樊后保, 黄玉梓, 袁颖红, 等. 2007. 森林生态系统碳循环对全球氮沉降的响应[J]. 生态学报, 27(7): 2997-3009.
樊后保, 黄玉梓, 袁颖红, 等. 2015. 森林生态系统碳循环对全球氮沉降的响应[J]. 生态学报, 27(7): 341-353.
方华, 莫江明. 2015. 氮沉降对森林凋落物分解的影响[J]. 生态学报,7(9): 357-366.
方运霆, 莫江明, GUNDERSEN P, et al. 2004. 森林土壤氮素转换及其对氮沉降的响应[J]. 生态学报, 24(7): 1523-1531.
黄雪夏, 唐晓红, 魏朝富, 等. 2007. 利用方式对紫色水稻土有机碳与颗粒态有机碳的影响[J]. 生态环境学报, 16(4): 1277-1281.
李德军, 莫江明, 方运霆, 等. 2003. 氮沉降对森林植物的影响[J]. 生态学报, 23(9): 1891-1900.
李德军, 莫江明, 方运霆, 等. 2015. 沉降对森林植物的影响[J]. 生态学报, 8(9): 211-220.
廖丹, 于东升, 赵永存, 等. 2015. 成都典型区水稻土有机碳组分构成及其影响因素研究[J]. 土壤学报, 52(3): 517-527.
林伟, 马红亮, 裴广廷, 等. 2016. 氮添加对亚热带森林土壤有机碳氮组分的影响[J]. 环境科学研究, 29(1): 67-76.
刘彩霞, 焦如珍, 董玉红, 等. 2015. 模拟氮沉降对杉木林土壤氮循环相关微生物的影响[J]. 林业科学, 51(4): 96-102.
龙凤玲, 李义勇, 方熊, 等. 2014. 大气CO2浓度上升和氮添加对南亚热带模拟森林生态系统土壤碳稳定性的影响[J]. 植物生态学报,38(10): 1053-1063.
马芬, 马红亮, 邱泓, 等. 2015. 水分状况与不同形态氮添加对亚热带森林土壤氮素净转化速率及 N2O排放的影响[J]. 应用生态学报,26(2): 379-387.
莫江明, 薛璟花, 方运霆. 2004. 鼎湖山主要森林植物凋落物分解及其对N沉降的响应[J]. 生态学报, 24(7): 1413-1420.
庞学勇, 刘世全, 刘庆, 等. 2003. 川西亚高山针叶林植物群落演替对土壤性质的影响[J]. 水土保持学报, 17(4): 42-45.
彭新华, 张斌, 赵其国. 2004. 土壤有机碳库与土壤结构稳定性关系的研究进展[J]. 土壤学报, 41(4): 618-623.
全权, 张震, 何念鹏, 等. 2015. 短期氮添加对东灵山三种森林土壤呼吸的影响[J]. 生态学杂志, 34(3): 797-804.
王晖, 莫江明, 鲁显楷, 等. 2008. 南亚热带森林土壤微生物量碳对氮沉降的响应[J]. 生态学报, 28(2): 470-478.
王晶苑, 张心昱, 温学发, 等. 2013. 氮沉降对森林土壤有机质和凋落物分解的影响及其微生物学机制[J]. 生态学报, 33(5): 1337-1346.
王绍强, 周成虎, 李克让, 等. 2000. 中国土壤有机碳库及空间分布特征分析[J]. 地理学报, 3(5): 533-544.
王绍强, 周成虎. 1999. 中国陆地土壤有机碳库的估算[J]. 地理研究,18(4): 349-356.
邬建红, 潘剑君, 葛序娟, 等. 2015. 不同农业利用方式土壤有机碳矿化及其与有机碳组分的关系[J]. 水土保持学报, 29(6): 178-183.
吴迪, 张蕊, 高升华, 等. 2015. 模拟氮沉降对长江中下游滩地杨树林土壤呼吸各组分的影响[J]. 生态学报, 35(3): 717-724.
吴金水. 2006. 土壤微生物生物量测定方法及其应用[M]. 北京: 气象出版社.
许凯, 徐钰, 张梦珊, 等. 2014. 氮添加对苏北沿海杨树人工林土壤活性有机碳库的影响[J]. 生态学杂志, 33(6): 1480-1486.
杨君珑, 李小伟. 2017. 宁夏不同草地类型土壤有机碳组分特征[J]. 生态环境学报, 26(1): 55-61.
游成铭, 胡中民, 郭群, 等. 2016. 氮添加对内蒙古温带典型草原生态系统碳交换的影响[J]. 生态学报, 36(8): 2142-2150.
张建华, 唐志尧, 沈海花, 等. 2017. 氮添加对北京东灵山地区灌丛土壤呼吸的影响[J]. 植物生态学报, 41(1): 81-94.
张俊华, 丁维新, 孟磊. 2010. 海南热带橡胶园土壤易氧化有机碳空间变异特征研究[J]. 生态环境学报, 19(11): 2563-2567.
张炜, 莫江明, 方运霆, 等. 2015. 氮沉降对森林土壤主要温室气体通量的影响[J]. 生态学报, 28(5): 431-441.
张英英, 蔡立群, 武均, 等. 2017. 不同耕作措施下陇中黄土高原旱作农田土壤活性有机碳组分及其与酶活性间的关系[J]. 干旱地区农业研究, 35(1): 1-7.
张忠启, 于东升, 潘剑君, 等. 2015. 红壤典型区不同类型土壤有机碳组分构成及空间分异研究[J]. 土壤, 47(2): 318-323.
