铜绿假单胞菌PA0745基因突变株的构建及鉴定
2018-06-07卢培林
卢培林,刘 利,熊 霞
(西南医科大学附属医院皮肤科,四川泸州 646000)
铜绿假单胞菌是一种革兰阴性需氧菌,普遍存在于土壤、水、空气和多种动物体内,具有较强的环境适应性和耐药性,可引起泌尿系统、呼吸系统和消化系统感染等,是慢性皮肤溃疡创面感染最常见的细菌。尤其在合并有基础疾病或免疫力低下时,如患有艾滋病、糖尿病和癌症等,病死率将大大增加。此外,由于铜绿假单胞菌耐药性强、耐药谱广,对多种抗菌药物表现出天然或获得性耐药,可供选择的有效抗菌药物很少。因此,从根本上寻找新的抗铜绿假单胞菌感染途径具有重大意义[1-2]。
研究显示,铜绿假单胞菌耐药性的产生与生物膜的形成息息相关。其中,临床分离的铜绿假单胞菌PA14细胞株可产生一种扩散信号分子(diffusible signal factor,DSF)类的小分子化合物顺式-2-癸烯酸(cis-2-decenoic acid,CDA),CDA可有效扩散PA14细胞株形成的生物膜,增强铜绿假单胞菌的致病性和耐药性。相关研究表明,烯酰辅酶A水合酶/异构酶是CDA合成过程的关键酶[3-6]。当PA0745基因功能受到抑制时,CDA合成受阻,铜绿假单胞菌菌膜扩散能力减弱,致病性下降。当外源性加入CDA时,菌膜运动能和细菌侵染力又得以恢复。另有研究表明,PA0745基因结构中有两个保守的谷氨酸残基对行使其催化功能具有重要意义,他们分别是α3上的Glu126和α4上的Glu146[7-9]。本研究利用同源重组的原理构建 PA0745基因点突变株E126A、E146A和基因缺失株ΔPA0745,通过生长曲线测定和细胞侵染实验初步验证了PA0745基因对铜绿假单胞菌致病性的影响。铜绿假单胞菌突变株的构建有助于开展铜绿假单胞菌功能组学研究和抗菌药物的开发。

表1 铜绿假单胞菌PA14突变株构建所用引物表
up:上游;down:下游
1 材料与方法
1.1材料 大肠埃希菌DH5α感受态细胞购自北京天根生化科技有限公司,S17-1由四川大学包锐研究员惠赠,铜绿假单胞菌PA14细胞株和穿梭质粒PEX18Gm均由四川大学王震玲教授惠赠。其中,PEX18Gm质粒含有负选择性标记基因SacB,含有庆大霉素(Gentamicin,Gm)抗性基因。
1.2主要试剂及引物 DNA聚合酶(Q5TMHot Start High-Fidelity DNA Polymerase)购自北京NEB公司,同源重组酶(ClonExpressTMⅡ One Step Cloning Kit)购自南京Vazyme公司,磷酸化酶(Blunting Kination Ligation Kit)购自日本TaKaRa公司,限制性内切酶(Dpn Ⅰ)购自美国Thermo Fisher Scientific公司,质粒提取试剂盒购自北京庄盟国际生物基因科技有限公司,抗生素购自北京天根生化科技有限公司,蛋白胨、酵母提取物购自英国Oxoid公司。引物合成和DNA测序由擎科生物科技有限公司完成,引物序列见表1。用Primer Priemier5.0软件,根据铜绿假单胞菌PA14株染色体上PA0745基因上下游序列,设计包含其上下游同源臂序列(800 bp)的引物、点突变引物和PA0745基因敲除引物。
1.3方法
1.3.1重组质粒PEX0745(PEX18+PA0745-up+PA0745+PA0745-down)的构建[10]
1.3.1.1制备线性化载体PEX18Gm 以PEX18Gm质粒为模板,以PEX18Gm-up和PEX18Gm-down为引物,用DNA聚合酶扩增质粒PEX18Gm,PCR反应条件如下:98 ℃ 30 s,98 ℃ 10 s,62 ℃ 25 s,72 ℃ 5 min,30个循环,72 ℃ 5 min。反应体系为25 μL,取5 μL PCR产物进行PCR鉴定。为了提高后期重组效率,需用限制性内切酶Dpn Ⅰ去除未线性化的环状质粒,条件如下:37 ℃ 2 h,80 ℃ 20 min。
1.3.1.2制备含目的基因上下游片段的PA0745-up+PA0745+PA0745-down 以PA14细胞株的DNA为模板,使用引物PEX0745-up和PEX0745-down扩增含上下游片段的PA0745-up+PA0745+PA0745-down,PCR反应条件如下:98 ℃ 30 s,98 ℃ 10 s,65 ℃ 30 s,72 ℃ 2 min,32个循环;72 ℃ 5 min。反应体系为25 μL,取5 μL进行PCR鉴定之后,再进行测序验证。
1.3.1.3利用重组酶连接目的片段和载体得重组质粒PEX0745 将克隆载体PEX18Gm和插入片段PA0745-up+PA0745+PA0745-down以摩尔比为1∶2的比例混合,利用重组酶进行重组反应得重组质粒PEX0745。反应条件如下:37 ℃ 30 min,4 ℃ 5 min。取5 μL进行PCR鉴定后,再进行测序验证。
1.3.2PA0745基因突变质粒E126A、 E146A和ΔPA0745的构建 以重组质粒PEX0745为模板,分别使用对应引物扩增出发生126位点突变、146位点突变和PA0745基因缺失的重组质粒E126A、E146A和ΔPA0745。PCR反应条件如下:98 ℃ 30 s,98 ℃ 10 s,62 ℃ 30 s,72 ℃ 5 min,30个循环;72 ℃ 5 min。取5 μL PCR产物进行PCR鉴定之后,进行测序验证。为了去除PCR产物中的质粒模板,需要对其进行Dpn Ⅰ酶切,条件如下:37 ℃ 2 h,80 ℃ 20 min。对酶切产物进行磷酸化及连接,反应条件如下:25 ℃ 5 min,16 ℃ 1 h。
1.3.3采用等位基因替换原理构建铜绿假单胞菌PA0745基因突变株 PEX18Gm为穿梭质粒,它可以通过结合转移的方法从大肠杆菌S17-1转移至铜绿假单胞菌,然后整合到其基因组中。PEX18Gm携带可以在大肠杆菌中用于质粒复制的复制子、Gm抗性基因和带有具有负选择性标记的基因SacB。基于铜绿假单胞菌可以利用柠檬酸作为碳源,而大肠杆菌不可以这一特点,利用仅含柠檬酸作为碳源的培养基VBMM作为第1次同源重组的筛选。另外,SacB基因的表达产物是果聚糖蔗糖酶,30 ℃条件下能催化蔗糖水解生成对大肠杆菌有致死作用的果聚糖。因此,SacB基因的表达会杀死生长在含有蔗糖培养基上的大肠杆菌,可以用做第2次同源重组的筛选。
分别将成功制备的重组质粒E126A、E146A和ΔPA0745转化到大肠杆菌S17-1感受态细胞中,后涂布于含有30 μg/mL Gm的LB琼脂板上,37 ℃培养过夜。
用LB培养基1∶1稀过夜培养的铜绿假单胞菌PA14细胞株,于42 ℃环境热阻断3~6 h,以体积比为1∶6的比例与对数期供体菌S7-1混合,用滤器过滤使细菌置于滤膜上,再把滤膜贴于LB琼脂板上,30 ℃孵箱孵育过夜。
1.3.4铜绿假单胞菌PA14突变株筛选鉴定 首先,利用含50 μg/mL Gm的VBMM琼脂糖平板进行第1次同源重组筛选;其次,利用含10%蔗糖的NSLB琼脂糖平板进行第2次同源重组筛选;最后,利用PIA琼脂糖平板分离铜绿假单胞菌,进行测序验证,得到铜绿假单胞菌PA14突变株E126A、E146A和ΔPA0745。
1.3.5铜绿假单胞菌PA0745基因突变株的生物学特性
1.3.5.1生长曲线测定 将铜绿假单胞菌PA14细胞株及其突变株E126A、E146A和ΔPA0745的过夜菌,分别按照1∶100的比例加入到3 mL新鲜的LB培养基和20% LB培养基中,37 ℃振荡培养2 h和3 h后,测OD600值,以后每0.5 h取样,测OD600值,绘制细菌体外生长曲线。
1.3.5.2细胞侵染实验 用含10% 胎牛血清(FBS)无抗生素的DMEM培养基调整对数期的铜绿假单胞菌PA14细胞株及其突变株E126A、E146A和ΔPA0745OD600为0.1,首先,细菌与巨噬细胞RAW264.7按照10∶1的感染复数(MOI)混合,37 ℃共孵育1 h。其次,培养基替换成含10% FBS和150 ng/mL Gm的DMEM培养基,37 ℃孵育1 h,用以除去细胞外铜绿假单胞菌。最后,磷酸缓冲盐溶液(PBS)清洗后,0.5% Triton X-100裂解细胞,梯度稀释后,涂布于LB琼脂糖平板上,次日对进入细胞内的细菌进行计数。

2 结 果
2.1重组载体的鉴定 以重组质粒PEX0745为模板,扩增出PA0745基因点突变株的质粒E126A、E146A和基因缺失株APA0745,并测序,测序结果与预期结果一致,见图1。

1:PA0745基因和其上下游800 bp片段;2:PA0745基因的上下游800 bp片段;3:pEX18Gm质粒线性化
图1 PCR产物琼脂糖凝胶电泳图谱
2.2PA14细胞株的PA0745基因突变株的特性 分别测试了PA14细胞株的PA0745基因突变株E126A、E146A和ΔPA0745在营养充足(LB)和营养缺陷(20% LB)的环境下的生长曲线。细菌在20% LB的生长条件下,8 h后细菌达到生长平台期,OD600为1.5左右,而在营养充足的LB生长条件下,8.5 h后细菌达到生长平台期,OD600为2.0左右。但不论细菌是在LB培养基中还是20% LB培养基中培养,PA14野生型菌株与突变株生长并没有差异,说明PA0745基因突变后不影响PA14细胞株的生长,见图2。

图2 铜绿假单胞菌PA14细胞株及突变株在LB(A)和20% LB(B)中生长曲线
2.3突变PA0745基因抑制铜绿假单胞菌细胞侵染能力比较 PA0745基因敲除后,铜绿假单胞菌侵入细胞的能力相较于野生型PA14细胞株下降了79.34%,具有催化活性的126、146位谷氨酸分别突变成丙氨酸后,相较于野生型PA14细胞株,细菌侵染能力也分别下降了29.51%及60.66%,见图3。
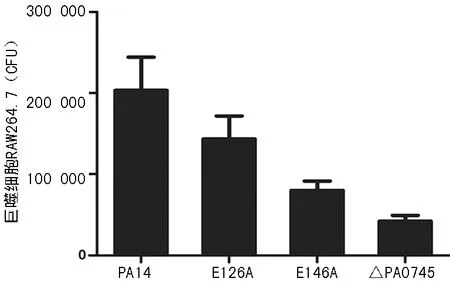
图3 铜绿假单胞菌PA14细胞株及其突变株侵染巨噬细胞RAW264.7能力比较
3 讨 论
慢性皮肤溃疡创面为铜绿假单胞菌的存活提供良好的“培养基”,使创面迁延不愈。而在近年相关研究中发现巨噬细胞在创面愈合过程中起总指挥的作用,当细菌大量繁殖扩增,使局部炎性反应及细胞因子的生物学效应减弱,影响巨噬细胞对抗炎和促进修复的调控。这为临床医生对慢性皮肤溃疡细菌感染的抗菌治疗提供理论依据[11]。
本研究通过同源重组的原理成功构建了铜绿假单胞菌PA0745基因突变株:E126A、E146A和ΔPA0745,通过生长曲线测定,各突变株与野生型PA14菌株之间并无明显差异,说明PA0745基因的功能缺失并不影响PA14细胞株的生长。通过巨噬细胞侵染实验考察PA0745基因对铜绿假单胞菌致病性的影响,发现相较于野生株PA14,PA0745基因各突变株的细胞侵染能力均出现了不同程度的下降。上述实验结果表明,以生物膜扩散信号分子形成过程中的关键酶为靶点,设计抗菌药物的可行性,由于它仅抑制细菌的致病性而不直接杀死细菌,这种抗菌药物将大大降低耐药菌株的产生。同时本实验从细菌侵染宿主方面证明了PA0745基因作为抗菌药物靶点的潜力,为开发针对PA0745基因靶点的新型抗菌药物提供理论基础[12]。
本研究针对铜绿假单胞菌生物膜扩散信号分子CDA生物合成的关键酶PA0745基因的功能展开了研究,在后续工作中,本课题组将通过对比分析铜绿假单胞菌野生型PA14和PA0745基因敲除株基因表达谱,深入研究CDA在铜绿假单胞菌内参与调控的下游信号通路,以期为以PA0745基因为靶点的小分子抑制剂的筛选提供理论基础和可行性分析。
[1]徐凤琴,黄松音,莫红平,等.sRNA phrs调控铜绿假单胞菌生物膜形成的功能研究[J].中华医院感染学杂志,2017,27(10):2169-2172.
[2]刘华之,侯良.皮肤创伤后铜绿假单胞菌生物膜感染的治疗效果研究[J].当代医学,2016,22(16):7-8.
[3]AMARI D T,MARQUES C N,DAVIES D G.The putative enoyl-coenzyme a hydratase dspi is required for production of the pseudomonas aeruginosa biofilm dispersion autoinducer cis-2-decenoic acid[J].J Bacteriol,2013,195(20):4600-4610.
[4]SEPEHR S,RAHMANIBADI A,BABAIENAIEJ H,et al.Unsaturated fatty acid,cis-2-decenoic acid,in combination with disinfectants or antibiotics removes pre-established biofilms formed by food-related bacteria[J].PLoS One,2014,9(7):e101677.
[5]JENNINGS J A,COURTNEY H S,HAGGARD W O.Cis-2-decenoic acid inhibits S.aureus growth and biofilm in vitro:a pilot study[J].Clin Orthop Relat Res,2012,470(10):2663-2670.
[6]RAHMANI B A,SEPEHR S,FALLAHI H,et al.Dissection of the cis-2-decenoic acid signaling network in Pseudomonas aeruginosa using microarray technique[J].Front Microbiol,2015,6:383.
[7] TAN D,CRABB W M,WHITMAN W B,et al.Crystal structure of DmdD,a crotonase superfamily enzyme that catalyzes the hydration and hydrolysis of methylthioacryloyl-CoA[J].PLoS One,2013,8(5):e63870.
[8]SPADARO F,SCOFFONE V C,CHIARELLI L R,et al.The crystal structure of burkholderia cenocepacia DfsA provides insights into substrate recognition and quorum sensing fatty acid biosynthesis[J].Biochemistry,2016,55(23):3241-3250.
[9]SRIVASTAVA S,CHAUDHARY S,THUKRAL L,et al.Unsaturated lipid assimilation by mycobacteria requires auxiliary cis-trans enoyl CoA isomerase[J].Chem Biol,2015,22(12):1577-1587.
[10]HMELO L R,BORLEE B R,ALMBLAD H,et al.Precision-engineering the Pseudomonas aeruginosa genome with two-step allelic exchange[J].Nat Protoc,2015,10(11):1820-1841.
[11]陈宏.巨噬细胞PPARγ对皮肤伤口愈合的作用研究[D].重庆:第三军医大学,2015.
[12]FONG J,YUAN M,JAKOBSEN T H,et al.Disulfide bond-containing ajoene analogues as novel quorum sensing inhibitors of pseudomonas aeruginosa.[J].J Med Chem,2017,60(1):215-227.
