HPC2蛋白在HPV16感染的宫颈癌细胞中调控作用初探*
2018-06-07杜海虹范凌晔毛熙光
刘 玲,杜海虹,范凌晔,聂 丹,毛熙光△
(1.西南医科大学附属医院妇产科,四川泸州 646000;2.四川省眉山市人民医院产科 620010)
宫颈癌是全球排名第3的妇女恶性肿瘤[1],人乳头瘤病毒(HPV)感染宫颈后,病毒基因整合到人宫颈上皮细胞,与其他促癌因素共同作用促成癌症的发生。E7基因是HPV的主要致癌基因之一,其编码的E7蛋白可使细胞周期失控而发生永生化。人多梳蛋白2 (HPC2)即染色盒同源物4(chrombox homolog 4,CBX4),在肿瘤细胞株和组织中表达上调,CBX4过表达与肿瘤分期和恶性程度密切相关[2],HPC2对靶基因的抑制作用的异常可能导致宫颈癌的发生[3]。为探索HPC2与宫颈癌发生的机制,本课题组研究了HPC2与HPV16的原癌基因E7之间的关系及其对细胞增殖、凋亡的作用,现报道如下。
1 材料与方法
1.1细胞与试剂 siha细胞(由西南医科大学附属医院中心实验室惠赠);Trizol、RNA提取试剂盒和cDNA试剂盒(北京天根生物科技有限公司),HPC2 siRNA(广州锐博生物科技有限公司),HPV16E7 siRNA(上海吉玛制药技术有限公司),lipofectamine 2000 reagent(美国Invitrogen公司),CBX4兔抗单克隆抗体(英国Abcam公司),β-actin兔抗单克隆抗体(美国Santa Cruz公司),CCK-8试剂盒(日本同仁公司),膜联蛋白V标记的异硫氰酸荧光素(Annexin V-FITC)细胞凋亡检测试剂盒(美国Thermo Fisher公司)。
1.2方法
1.2.1实验分组 将siha细胞分为4个组:空白对照组、NControl siRNA组(对照组)、HPC2 siRNA组和HPV16E7 siRNA组。空白对照组为未转染任何序列RNA组,NControl siRNA组为转染阴性序列对照组,HPC2 siRNA组为沉默HPC2组,HPV16E7 siRNA组为沉默HPV16E7组。
1.2.2细胞转染 每个6孔培养板中接种4×105个对数生长期的siha细胞,培养24 h,细胞融合达到60%时开始实验。操作步骤按照美国Invitrogen公司提供的操作说明。倒置荧光显微镜下观察转染24 h后NControl siRNA-CY3转染组红色荧光情况。HPC2 siRNA(广州市锐博生物科技有限公司合成),靶序列:5′-GGT AGA ATG TCC AGA TGA A-3′,正义链:5′-GGU AGA AUG UCC AGA UGA AdTdT-3′,反义链:3′-dTdTC CAU CUU ACA GGU CUA CUU-5′。HPV16E7 siRNA(上海吉玛公司合成),5′-GGA CAG AGC CCA UUA CAA UTT AUU GUA AUG GGC UCU GUC CTT-3′。
1.2.3荧光定量PCR 提取细胞总RNA参照北京天根公司提供的说明书操作,引物由上海生工生物工程公司合成,引物序列如下。甘油醛-3-磷酸脱氢酶(GAPDH)引物序列:上游5′-CCC CAT ACA CAG TGT TAG CC-3′,下游5′-GAG TGA TTT TCC CGT CC-3′;HPC2引物序列:上游5′-ACC ACA GTC CAT GCC ATC AC-3′,下游5′-TCC ACC ACC CTG TTG CTG TA-3′;HPV16E7引物序列:上游5′-TGC AAC CAG CAA CTG AT-3′,下游5′-TGC AAC CAG CAA CTG AT-3′。DNA扩增条件为94 ℃预变性3 min;94 ℃变性30 s,50 ℃退火1 min,72 ℃延伸1 min,循环35次;终延伸72 ℃ 5 min。荧光定量PCR反应条件为96 ℃预变性5 min;96 ℃变性30 s,57 ℃退火30 s,72 ℃延伸30,循环40次;终延伸72 ℃ 10 min。每组设3个复孔,记录Ct值,取平均值。△Ct值=目的基因Ct值-内参基因的Ct 值;△△Ct值=实验组基因的△Ct值-空白对照组基因的△Ct 值;2-△△Ct值=实验组基因/空白组基因(倍数)。
1.2.4Western blot 提取总蛋白并测其浓度,制胶,正丁醇封闭20~30 min,上样,电泳,转膜,5%脱脂奶粉封闭,置于摇床上摇2 h,封闭液稀释一抗HPC2(1∶400)、β-actin(1∶1 000)孵育;封闭液稀释兔二抗(1∶1 000)孵育。化学发光后凝胶成像系统成像,保存图片。
1.2.5CCK-8试验 每组设5个复孔,将96孔板放细胞培养箱24 h后转染siRNA,细胞转染1~7 d时间点每孔加入10 μL CCK-8 溶液,放进细胞培养箱继续培养,于2 h时间点用酶标仪测定在450 nm处的吸光度(OD)。
1.2.6流式细胞技术 siha细胞接种于25 cm2细胞培养瓶,24 h后行细胞转染,转染48 h收集细胞,1 000 r/min离心5 min;弃上清液,加入195 μL 结合缓冲液轻轻重悬细胞,加入5 μL Annexin V-FITC轻轻混匀,避光室温孵育15 min;离心后弃上清液,加入190 μL Annexin V-FITC 结合缓冲液轻轻重悬细胞,10 μL碘化丙啶(PI)溶液,轻轻混匀,避光室温放置20 min,冰浴;上机检测。

2 结 果
2.1siha细胞HPV16E7基因沉默后HPC2基因的表达 siha细胞转染HPV16E7 siRNA 48 h后,HPC2 mRNA表达水平较对照组下调75%左右,差异有统计学意义(P<0.01),见图1。

a:P<0.01。
图1 3组HPC2 mRNA(2-△△Ct值柱状图)在siha细胞中的表达
2.2HPV16E7基因抑制后对siha细胞中HPC2蛋白表达的影响 siha细胞转染HPV16E7 siRNA 48 h后,HPC2蛋白水平下调,与HPC2 siRNA转染效果相当,见图2。

1,4:空白对照组;2,5:对照;3:HPC2 siRNA组;6:HPV16E7 siRNA组
图2 6组HPC2蛋白在siha细胞中的表达
2.3HPC2基因抑制后对siha细胞增殖的影响 随着时间延长,OD值逐渐增加。与其他各组细胞相比,转染HPC2 siRNA的siha 细胞OD值自第3天开始增幅降低,第7天各组细胞OD值分别为:空白对照组2.08±0.07;对照组2.00±0.09;HPC2 siRNA组0.92±0.10。HPC2 siRNA组较对照组显著降低(t=20.403,P<0.01),见图3。
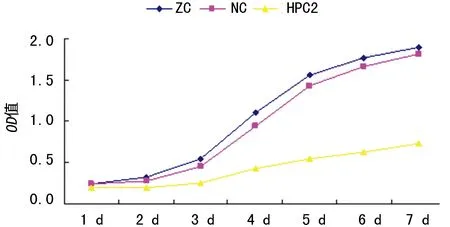
ZC:空白对照组;NC:对照组;HPC2:HPC2 siRNA组
图3 3组siha细胞OD值变化
2.4HPC2基因抑制后对siha细胞凋亡的影响 转染48 h后,HPC2 siRNA组早期凋亡和晚期凋亡比例为6.38%,HPV16E7 siRNA组为7.03%,对照组为3.94%,空白对照组为0.21%;HPC2 siRNA组转染后细胞凋亡比例较对照组增加2.44%,HPV16E7 siRNA组转染后细胞凋亡比例较对照组增加3.09%,见图4。

图4 4组 siha细胞凋亡率
3 讨 论
HPC2作为PcG家族重要成员,是基因转录抑制因子,与B细胞特异性莫洛尼鼠白血病病毒插入位点-1(BMI-1)、RING1、HPH3 3种 PcG蛋白及非PcG蛋白C-末端结合蛋白(CtBP)和转录因子E2F相互作用形成HPC-HPH蛋白复合物来执行功能[4]。HPC2可能抑制原癌基因的转录而参与调控肿瘤形成、细胞凋亡及细胞周期等[5]。HPC2既是PcG蛋白又是SUMO E3连接酶,其可通过SUMO化修饰调节KDM5B活性,抑制细胞周期和DNA修复基因的表达[5];CBX4通过募集HCAC3到RUNX2启动子抑制RUNX2的表达[6]。人类癌细胞系和正常组织中HPC2的表达水平差异较大,在Burkitt淋巴瘤和肺癌中几乎检测不到HPC2转录子,而HPC2在其他细胞系中均呈高表达状态[5]。HPC2基因突变或表达异常与肿瘤的发生可能存在密切联系[7-8],CBX4通过抑制RUNX2的表达抑制结肠癌的转移[6,9],CBX4 通过控制HIF-1α蛋白在肿瘤血管生成中起着关键作用,作为SUMO E3泛素连接酶促进肝癌的进展和转移[7-8],敲除CBX4导致了HIF-1α靶基因的下调表达,导致在常氧条件下骨肉瘤细胞生长和细胞存活率的显著抑制[2]。既往研究证实PC家族中HPC2是惟一具有SUMO E3活性的成员[10],在细胞干性、细胞衰老/胚胎发育、细胞周期、DNA损伤应答及肿瘤的发生等方面均有调控作用。
羧基端是HPC2蛋白发挥抑制功能必不可少的结构域,若其发生突变会导致其抑制靶基因转录的功能丧失。但其可与内源性的HPC2 蛋白竞争结合PcG蛋白反应元件,从而干扰正常内源性HPC2蛋白的功能。HPC2的SUMO化作用能改变蛋白构型,进而改变蛋白功能。有研究检测到人宫颈癌HPC2基因的C端的保守序列的突变,其与转录抑制物CtBP结合的能力丧失,抑制靶基因转录的作用降低,细胞恶性分化,诱发宫颈癌[3]。以上文献报道基于对HPC2结构水平上的研究,而本实验从定量上探讨HPC2与E7基因的关系,结果显示:E7基因沉默后,HPC2无论是在基因水平还是在蛋白水平均呈现下调表达,推测E7蛋白可能调控PcG蛋白HPC2的表达。
Notch信号转导途径的变化与某些肿瘤的发生、发展密切相关。HPV相关蛋白E6/E7介导宫颈癌细胞Notch信号通路的异常上调,二者在维持肿瘤的生长及异型性有协同作用。此外,Fas蛋白家族介导Notch1抑制E6/E7的表达。Notch信号途径参与了细胞增殖、凋亡与分化等过程,参与肿瘤的发生[11]。关键转录因子CBF-1/RBP-Jk介导了经典的Notch信号途径,而HPC2参与了对RBP-Jk的调控[12]。本研究结果提示E7调控HPC2的表达,由此推测E7可能通过调控HPC2的表达来调控RBP-Jk介导的经典Notch通路,Notch通路的异常调控,最终导致宫颈癌的发生。本研究发现抑制HPC2与抑制E7基因后对宫颈癌细胞增殖和凋亡的影响相似,结合E7与HPC2基因之间的关系,推测E7基因可能通过调控HPC2来参与细胞的增殖凋亡过程,从而参与宫颈癌的发生。但其具体通过何种途径何种机制来参与,还需要更进一步的研究来证实。
[1]FERLAY J,SOERJOMATARAM I,DIKSHIT R,et al.Cancer incidence and mortality worldwide:sources,methods and major patterns in GLOBOCAN 2012[J].Int J Cancer,2015,136(5):359-386.
[2]LIN F M,KUMAR S,REN J,et al.SUM Oylation of HP1α supports association with ncRNA to define responsiveness of breast cancer cells to chemotherapy[J].Oncotarget,2016,7(21):30336-30349.
[3]秦鸿雁,王林,杨红等.HPC2基因与宫颈癌关系的研究 [J].第四军医大学学报,2004,25(2):166-168.
[4]SEWALT R G,LACHNER M,VARGAS M,et al.Selective interactions between vertebrate polycomb homologs and the SUV39H1 HMTase suggest histone H3-K9 methylation to contribute to chromosomal targeting of Polycomb-group proteins [J].Mol Cell Biol,2002,22(15):5539-5553.
[5]BUENO M T,RICHARD S,REN J,et al.SUMOylation negatively modulates target gene occupancy of the KDM5B,a histone lysine demethylase[J].Epigenetics,2013,8(11):1162-1175.
[6]WANG X,KANG T.Role of CBX4 in the colorectal carcinoma metastasis-response[J].Cancer Res,2017,77(9):2550-2551.
[7]MEI Z,JIAO H,WANG W,et al.Polycomb chromobox 4 enhances migration and pulmonary metastasis of hepatocellular carcinoma cell line MHCC97L[J].Sci China Life Sci,2014,57(6):610-617.
[8]LI J,XU Y,LONG X D,et al.Cbx4 governs HIF-1α to potentiate angiogenesis of hepatocellular carcinoma by its SUMO E3 ligase activity[J].Cancer Cell,2014,25(1):118-131.
[9]SANCISI V,CIARROCCHI A.Role of CBX4 in the colorectal carcinoma metastasis-letter[J].Cancer Res,2017,77(9):2548-2549.
[10]AGRAWAL N,BANERJEE R.Human polycomb 2 protein is a SUMO E3 ligase and alleviates substrate-induced inhibition of cystathionine beta-synthase sumoylation[J].PLoS One,2008,3(12):e4032.
[11]YUAN X,WU H,XU H,et al.Notch signaling:an emerging therapeutic target for cancer treatment[J].Cancer Lett,2015,369(1):20-27.
[12]QIN H,DU D,ZHU Y,et al.The PcG protein HPC2 inhibits RBP-J-mediated transcription by interacting with LIM protein KyoT2[J].FEBS Lett,2005,579(5):1220-1226.
