巨菌草不同生长时期根、茎、叶内生细菌组成的多样性及差异
2018-06-06林标声宋昭昭张丽丽范锦琳林占熺
林标声, 宋昭昭, 张丽丽, 范锦琳, 林占熺
(1.福建农林大学生命科学学院,福建 福州 350002;2.龙岩学院生命科学学院,福建 龙岩 364012;3.国家菌草工程技术研究中心,福建 福州 350002)
植物内生细菌是定殖于植物组织内部,对植物生长、病害防治起到促进作用的一类生物[1-2].植物内生细菌的研究起源于20世纪40年代末50年代初[3-4],研究者发现在许多植物的不同组织内都存在着不同种类的内生细菌[5-8],如禾本科植物的玉米、水稻、小麦、甘蔗等,并从其苗期、穗期的根、茎、叶等部位分离得到了芽孢杆菌属(Bacillus)、肠杆菌属(Enterobacter)、黄单胞菌属(Xanthomonas)、假单胞菌属(Pseudomonadaceae)、变形杆菌属(Proteus)、细杆菌属(Microbacterium)、根瘤菌属(Rhizobium)及固氮菌属(Azotobacter)等内生细菌.这些内生细菌种类繁多,生物多样性丰富,组成及群落结构因植物种类、组织器官及生长时期的不同而差异较大[9-11].研究表明,植物内生细菌的菌群密度、数量与植物种类、生长环境、发育时期和组织器官均有关系,随着植物的生长,植物内生菌群的密度和数量均表现出较为明显的增长趋势,这种增长趋势对促进植物生长、代谢产物合成以及增强植物抗病性等方面具有重要作用[12-14].因此了解植物内生细菌组成及多样性对优化植物生长状态、维持生态系统的平衡和生物多样性具有重要的意义.
巨菌草(Pennisetumsp.)是2005—2007年由福建农林大学菌草研究所从南非莱索托引进的品种,隶属于狼尾草属,多年生,直立,丛生,根系发达,抗逆性强,产草量高.巨菌草能在中国的西藏、内蒙古、青海、宁夏等土地贫瘠的沙漠种植生长,种植后沙地的有机质、微生物数量明显增加,酶活性明显增强,固沙效果明显[15].同时,巨菌草的总氮含量超过了其从土壤和肥料中摄取的总氮量,很可能是巨菌草内生固氮细菌在起作用.近年来,许多研究者对甘蔗、玉米、水稻等禾本科植物中发现的多种具有固氮功能的内生细菌进行了广泛的研究,成为联合固氮研究领域的热点[16-17].巨菌草内生固氮细菌资源的发掘与利用,在促进植物生长、增强植物抗逆性、提高植物对土壤营养物质的利用率、改善荒漠生态环境等方面均具有重要的研究价值,可为将来大力推广荒坡地种植巨菌草,提高菌草生物质产量,开拓内生菌生物肥料应用提供新思路、新途径[18].因此,本试验以巨菌草不同生长时期根、茎、叶为研究对象,采用Illumina Miseq第2代测序技术深入分析其内生细菌的种类、分布及变化规律,旨在更加全面、准确地的反映巨菌草不同生长时期内生菌群的组成和差异,探寻、丰富和完善巨菌草的内生细菌种质资源,为归纳和总结巨菌草内生细菌在宿主内分布、抗逆及促生长等作用提供理论基础.
1 材料与方法
1.1 样本的采集及处理

表1 各采样时期的环境指标Table 1 Environmental parameters at different sampling times
2016年8月至12月,在福建农林大学旗山校区菌草研究所试验基地同一试验地采集5个不同生长时期的巨菌草根、茎、叶,采用不同分布多位点采集,采集后立即清洗干净,混合均匀后装入无菌封口塑料袋,共15个样本.各采样时期的环境因素(空气相对湿度、温度和土壤pH)见表1.
1.2 样本的表面消毒
挑选各样本中无病变、品相良好的巨菌草根、茎、叶部位3~5 g,先用流水冲洗1~2 h,再依次采用75%乙醇、2%次氯酸钠、无菌水洗涤.其中先用75%乙醇浸泡60 s,然后根、茎、叶分别用2%次氯酸钠浸泡20 min、9 min、90 s进行表面灭菌,最后用无菌水冲洗4次.采用稀释涂布法确认待分离材料是否消毒彻底,即将最后一次的无菌水洗涤液涂布于LB培养基中,30 ℃培养1~2 d,以无菌在LB培养基上生长为消毒合格.
1.3 样本细菌16S V3~V4可变区的PCR扩增和高通量测序
采用OMEGA试剂盒提取,具体方法参照试剂盒说明书.PCR扩增选择长度约为450 bp的细菌作为目的扩增片段,进行后续高通量测序.引物序列为:前引物ACTCCTACGGGAGGCAGCA,后引物GGACTACHVGGGTWTCTAAT.PCR反应体系(25 μL):5×反应缓冲液5 μL,5×GC缓冲液 5 μL,dNTP(100 mmol·L-1)2 μL,正向引物(10 umol·L-1)1 μL,反向引物(10 umol·L-1)1 μL,Q5高保真度DNA聚合酶0.25 μL,DNA模板2 μL,ddH2O 8.75 μL;PCR反应条件为:98 ℃变性2 min,98 ℃变性15 s,55 ℃退火30 s,72 ℃延伸30 s,扩增25~30个循环,最后于72 ℃延伸5 min.扩增结果进行2%琼脂糖凝胶电泳,切取目的片段后用Axygen凝胶回收试剂盒回收目的片段,随后交由上海派森诺生物科技股份有限公司使用Illumina Miseq高通量测序平台进行序列测定和分析.
1.4 测序结果的数据处理与分析
首先,用Qiime 1.17软件去杂、拼接,去除Illumina Miseq测得的质量不好的序列,再通过嵌合体序列处理去杂后的数据,确定最终的有效序列.应用Usearch软件,按照97%相似度的CD-HIT分类方法对有效序列进行OUT聚类,以丰度最大的序列为代表序列,统计各个样本OUT的量及每个OUT所含的有效序列数目,分析各样本在各分类水平上菌群组成及多样性[19].根据各样本中在属水平上的丰度信息,从物种和样本两个层面进行聚类,采用R软件绘图,分析各样本组间的相似性.
采用Canoco for Windows 4.5软件分析环境因子对各样本细菌群落结构的影响,以属水平上丰度最高的前20个属作为物种依据进行典范对应分析(canonical correspondence analysis, CCA).
2 结果与分析
2.1 DNA提取与16S rDNA扩增
从各样本中提取的DNA和16S rDNA V3~V4可变区扩增结果如图1和图2所示.图中PCR产物目标条带正确,浓度满足建库需要.
1~3:苗期根、茎、叶;4~6:分蘖期根、茎、叶;7~9:拔节期根、茎、叶;10~12:成熟期根、茎、叶;13~15:衰亡期根、茎、叶;M:Marker DL 15 000 bp.图1 各样本DNA电泳结果Fig.1 Electropherogram of PCR amplified samples

各样本情况详见图1.图2 提取各样本16S rDNA V3~V4可变区扩增结果图Fig.2 Electropherogram of PCR amplified 16S rDNA V3 ~V4 variable range of samples
2.2 测序结果的质量分析
巨菌草不同生长时期的15个样本,得到二、三和四万多条有效序列的样本数分别是3、10和2个(表2),其在不同生长时期操作分类单元(operational taxonomic unit, OTU) 的数量均为根>茎>叶,菌群细菌多样性指数ACE、Chao1、Shannon、Simpson也均为根>茎>叶,表明巨菌草在不同生长时期细菌多样性和丰度均是根>茎>叶.从有效序列长度图可得(图3),序列主要分布在400~450 bp.各样本的稀释度曲线如图4所示,15条曲线均趋向平坦,不同生长时期的根、茎、叶曲线均较为集中地聚类在一起,表明测序数据量合理,大多数细菌类群包含在了各样本文库中,能够比较真实地反应样本的细菌群落多样性[20].
2.3 各样本物种丰度及群落结构
从门的分类水平看(图5),巨菌草不同生长时期根、茎、叶的内生细菌均以蓝藻菌门和变形菌门为主,其他丰度较高的种类还包括放线菌门(Actinobacteria)、拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)和绿弯菌门(Chloroflexi);拔节期菌群种类多于其他时期,且同一生长时期菌群种类根>茎>叶,根部菌群的种类及丰度明显高于茎部和叶部.如分蘖期根部(序号4)出现了2.9%的绿弯菌门(Chloroflexi),但茎、叶中均未发现;成熟期根部(序号10)出现了大量放线菌门(Actinobacteria),其菌群丰度达到28.2%,但同时期茎部的丰度仅为0.8%,叶中未发现.此外,在根部分布较多的拟杆菌门(Bacteroidetes)(丰度0.2%~4.2%),在茎中较少(丰度0.1%),在叶中未发现;在根、茎中发现少量的硬壁菌门(Firmicutes)(丰度0.2%~1.9%)、酸杆菌门(Acidobacteria)(丰度0.1%~0.3%),在叶中也未发现;根中还发现了微量疣微菌门(Verrucomicrobia)、OP11、TM7、浮霉菌门(Planctomycetes)(丰度均为0.1%左右),但在茎、叶中均未发现.
在属的水平上,取所有样本中平均相对丰度前20的菌群,剩余所有分类单元归为“Other”再作图,分析巨菌草在不同生长时期的细菌菌群分布和变化情况(图6).结果表明,根的细菌菌群种类和丰度均要明显的多于茎、叶,且根>茎>叶.各个样本中,丰度较高的属为罗尔斯顿菌属(Ralstonia)、乳球菌属(Lactococcus)、农杆菌属(Agrobacterium)、伯克氏菌属(Burkholderia)、鞘脂单胞菌属(Sphingomonas)、细杆菌属(Microbacterium)、甲基杆菌属(Methylobacterium)和伦茨氏菌属(Lentzea),其中罗尔斯顿菌属(Ralstonia)和乳球菌属(Lactococcus)为核心属,始终存在于各个样本中,丰度比例分别为1.8%~44.3%和1.6%~24.8%.此外,各个样本中还存在着大量的“Other”其他菌属,其中拔节期(序号7~9)的丰度最高,表明此时期巨菌草快速生长,菌群种类比较复杂;衰亡期(序号13~15)“Other”其他菌属的丰度较其他时期略高,原因可能是巨菌草生长时期延长,内生侵入菌群增多,另外的可能是植物生长末期抗性降低,大量外部细菌菌群更易侵染植物,导致其内生菌群种类增多[21],如禾本科植物的白叶枯黄致病菌黄单胞菌属在衰亡期根茎、叶、中出现,但在其他时期未发现.
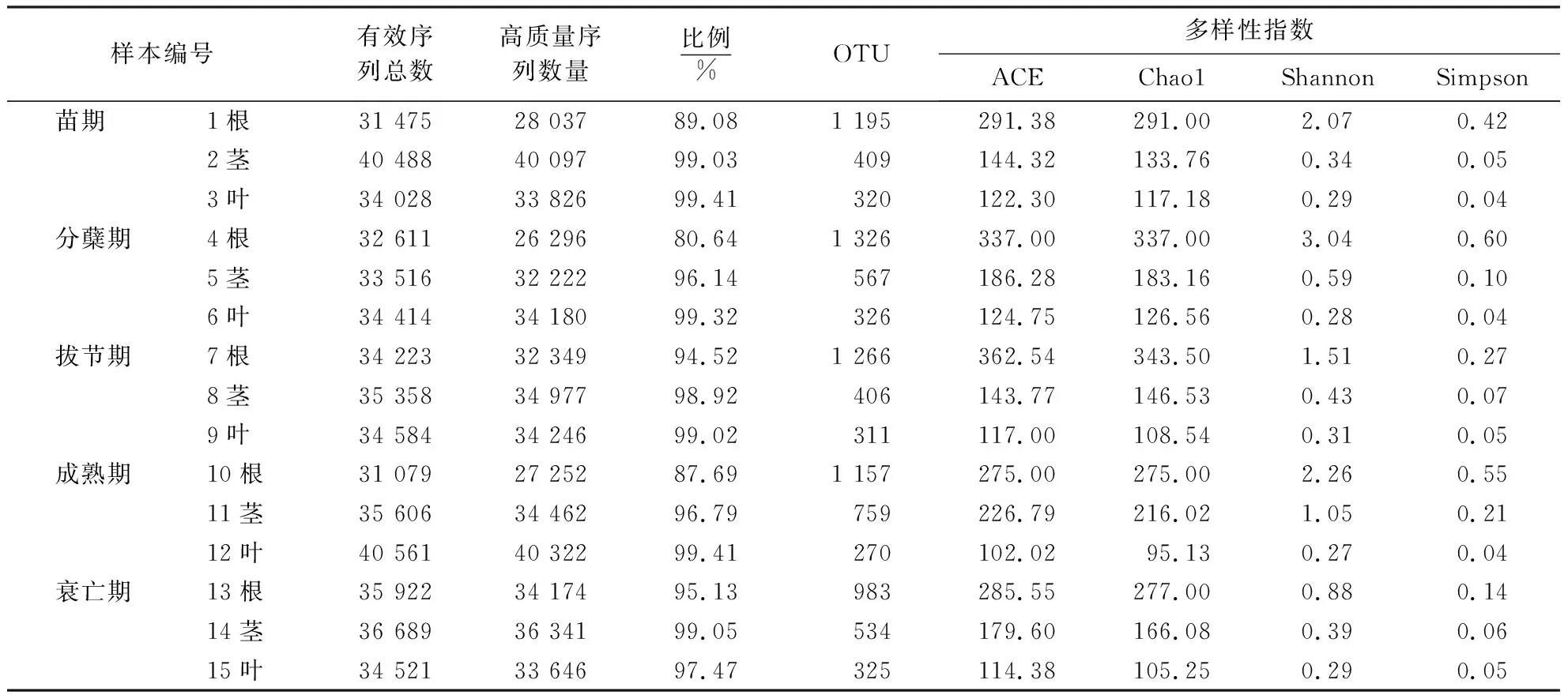
表2 各样本的测序数据、OTU及多样性指数统计Table 2 Sequencing data, OTU and diversity index statistics of each sample

图3 各样本序列长度分布图Fig.3 Length distribution of each sample

各样本情况详见图1.图5 各样本在门分类水平上主要细菌类群比较Fig.5 Comparison of major bacterial class in each sample on phylum level
基于GraPhlAn的样本总体分类绘制等级树图,相对丰度前20位的分类单元已在图中显示(图7),结果表明:门水平上丰度较高的是蓝藻菌门、变形菌门、拟杆菌门和放线菌门;属水平上丰度较高的是农杆菌属和伦茨氏菌属.所得的结果与上述分类统计结果一致.
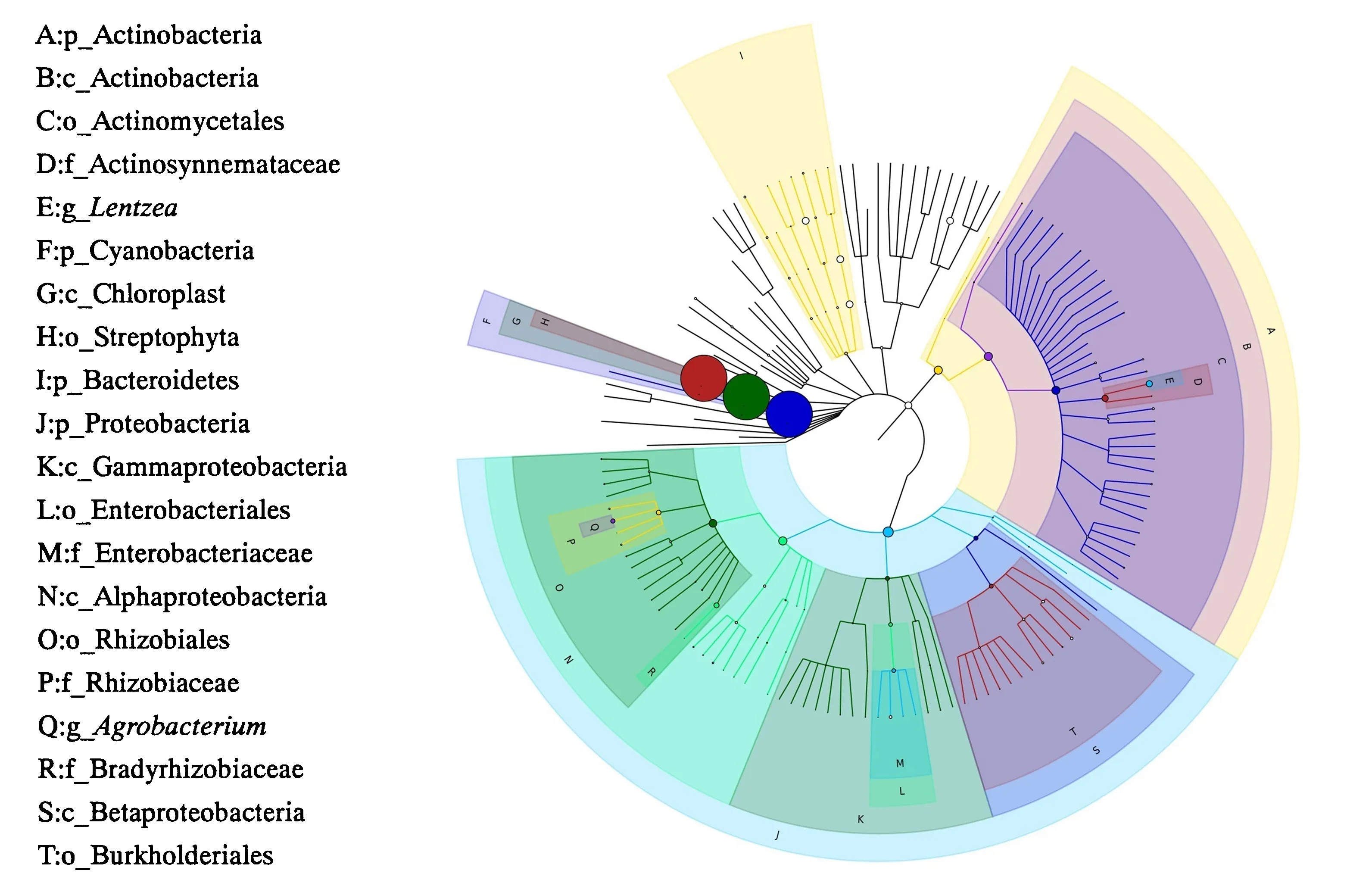
图7 基于GraPhlAn的样本总体分类等级树图Fig.7 Hierarchy tree classification of all samples based on GraPhlAn
2.4 各样本组间的相似性分析
使用R软件,对丰度前50的属进行聚类分析得Heatmap(图8).如图可得,同一部位来源的样本,如来自根部(序号1、4、7、10、13)和叶部(序号3、6、9、12、15)的菌群可较好地聚类在一起,相互间无交叉;茎部样本(序号2、5、8、11、14)较分散,其内部和根部、叶部样本间交叉聚类,表明茎部细菌菌群可能有部分来源于根部和叶部.根据图中颜色梯度的变化可得,各类细菌菌群在根部样本的颜色明显深于茎部、叶部,表明根部各类菌群的丰度及表达量均明显高于茎部、叶部,且其在不同时期的菌群组成不同,变化较复杂;叶部各类菌群的颜色组成及变化基本一致,表明来自叶部的细菌菌群组成具较高相似性.
通过R软件,对属水平的群落组成结构进行主成分分析(图9),其第1主成分和第2主成分的贡献率分别为80.78%和13.79%,除了样本1、4和10外,其他样本的遗传距离较为接近,所得结果基本与Heatmap结果吻合.

各样本情况详见图1.图8 各样本中属水平细菌菌群Heatmap分析Fig.8 Heatmap analysis of microbial community under genus level in each sample

各样本情况详见图1.图9 各样本中属水平细菌菌群PCA分析Fig.9 PCA analysis of microbial community under genus level in each sample
2.5 环境因素对各样本细菌菌群组成的影响
对属水平丰度前20的属进行CCA分析,研究空气湿度、温度和土壤pH对样本细菌菌属的变化情况(图10).结果表明:轴1和轴2各样本细菌属的变化分别解释了36.8%和30.6%的变异,即所有的环境因素对总变异共有66.8%的解释度.图中,丰度前20的细菌属的丰度变化与温度有关,其次是湿度和pH.湿度和pH变量箭头之间的夹角较小,表明二者对细菌属的变化可能具有协同效应[22].20个属中有11个属与温度正相关,包括农杆菌属、伯克氏菌属等具有固氮作用的菌群.
3 小结
本试验首次采用了高通量测序技术研究了巨菌草内生细菌菌群的变化情况,表明巨菌草不同生长时期根、茎、叶的细菌组成及丰度存在较大的差异,其15个样本中,得到二、三和四万多条有效序列的样本数分别是3、10和2个,主要分布在400~450 bp.从门到属的水平上看,巨菌草内生菌群组成均为拔节期较高,在同一生长时期则为根>茎>叶;蓝藻菌门、变形菌门和罗尔斯顿菌属、乳球菌属分别是巨菌草所有分离样本的主要门类和核心属类.在门水平上,各样本中丰度较高的还有放线菌门,而蓝藻菌门和放线菌门是探讨生物进化、挖掘农用抗生素菌群的重要种质资源来源[23].在属水平上,根部来源的各样本中均含有较高丰度的农杆菌属、伯克氏菌属,这两种菌均属于变形菌门,其在氮肥开发与利用、农业生物基因工程方面具有广泛的用途[24-25],是具有重要农业应用价值的固氮细菌种质资源.因此,本试验中所得的巨菌草在不同生长时期的菌群丰度变化可为挖掘巨菌草具有特定应用价值的菌群资源指明方向.
本试验中,Heatmap聚类和主成分分析表明,巨菌草不同生长时期根部、叶部来源的细菌菌群组成较为接近,茎部来源菌群与根部、叶部有交叉,表明其菌群组成可能有部分来源于根部和叶部.因此,下一步的研究将采用特定标记的菌群在巨菌草中进行定殖试验,观察根部和叶部的细菌菌群是通过何种特定的运输途径转移到茎部[27].此外,巨菌草不同生长时期根、茎、叶样本之间的细菌菌群组成及丰度差异较大,变化情况不同,推测巨菌草从苗期到衰亡期内生菌群的组成可能受到空气湿度、温度、土壤等环境因素的影响.CCA分析表明,温度是影响巨菌草内生固氮菌菌群变化的主要因素,其次是湿度和pH.丰度最高的20个属种有11个属与温度影响呈正相关,其中包括促进植物生长农杆菌属、伯克氏菌属等具有固氮作用的菌群,表明较高的温度条件下巨菌草内生固氮菌群增多,联合固氮作用增强,有利于植物的生长.
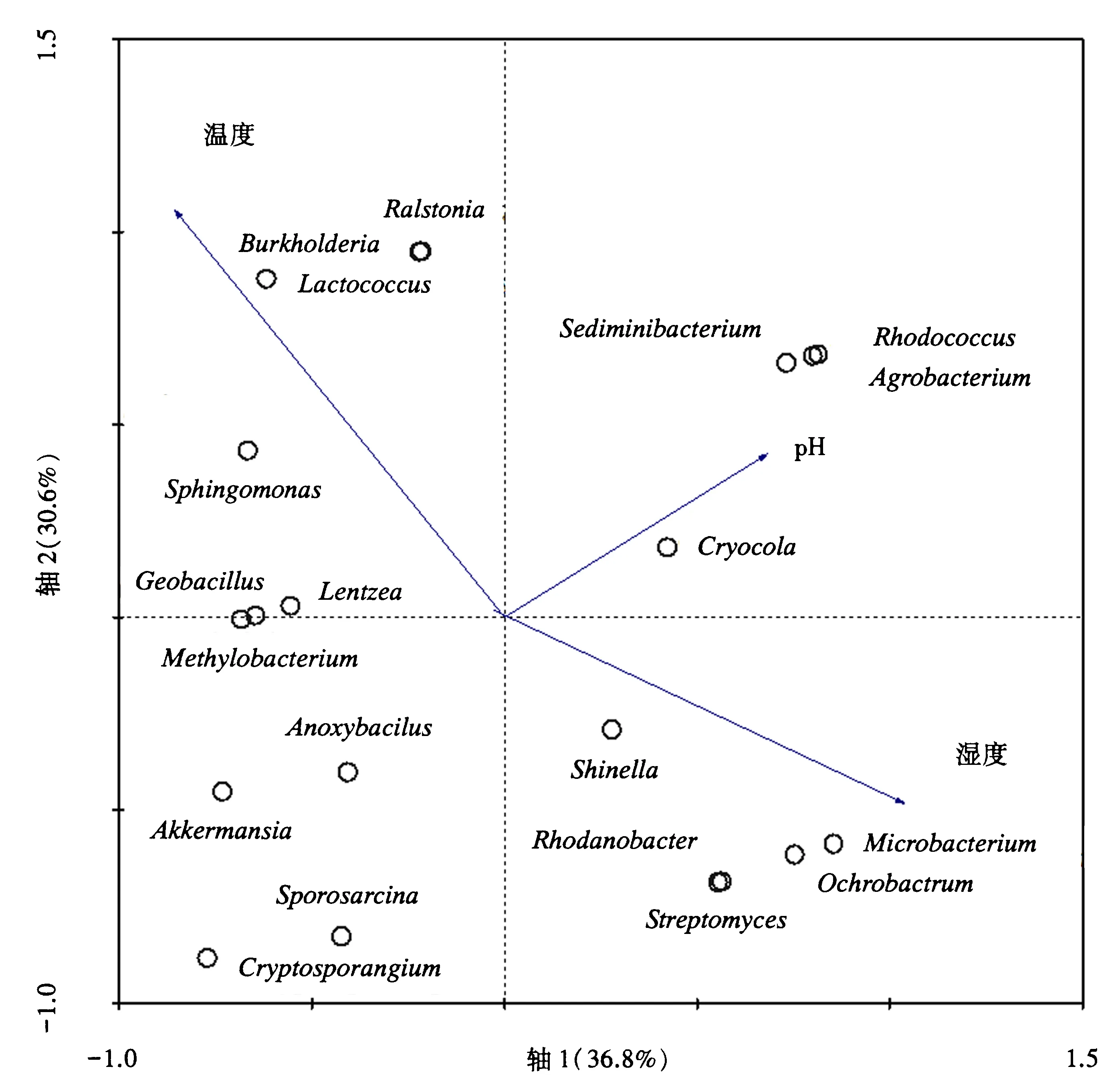
图10 环境因素对各样本细菌菌群组成影响的CCA分析Fig.10 CCA analysis on the environmental factors influence of microbial community composition in each sample
综上所述,较高温度条件下,巨菌草生长到拔节期,其根部的内生菌群种类及丰度最多,有利于细菌种质资源的挖掘与开发.本试验结果可为进一步挖掘巨菌草可培养内生细菌资源,建立种质资源库,开发和利用根瘤菌、固氮菌、产抗生素细菌等农用微生物菌剂提供基础依据[27].
[1] LIAQAT F, ELTEM R. Identification and characterization of endophytic bacteria isolated from in vitro cultures of peach and pear rootstocks[J]. Biotechnology, 2016,120(6):1-8.
[2] 丁文珺,王珊珊,任婧祺,等.植物内生菌研究进展[J].生物技术进展,2015,5(6):425-428.
[3] TERVET W I, HOLLIS J P. Bacteria in the storage organs of healthy plants[J]. Phytopathology, 1948,38(12):960-967.
[4] HOLLIS J P. Bacteria in the healthy potato tissue[J]. Phytopathology, 1951,41:350-366.
[5] SZILAGYIZECCHIN V J, IKEDA A C, HUNGRIA M, et al. Identification and characterization of endophytic bacteria from corn (ZeamaysL.) roots with biotechnological potential in agriculture[J]. Amb Express, 2014(4):1-9.
[6] 王伟平,李南南,黎妮,等.基于微生物分离培养的超级杂交水稻深两优5814种子内生细菌多样性研究[J].杂交水稻,2016(4):61-66.
[7] PAN D, MIONETTO A, TISCORNIA S, et al. Endophytic bacteria from wheat grain as biocontrol agents of Fusarium graminearum, and deoxynivalenol production in wheat[J]. Mycotoxin Research, 2015,31(3):137-143.
[8] 王丽,张剑亮,王继华,等.甘蔗茎部内生细菌的分离及分子鉴定[J].热带作物学报,2013,34(11):2 227-2 232.
[9] 杨文琦,龙宣杞,崔卫东.玉米青贮中细菌多样性分析[J].新疆农业科学,2013,50(8):1 424-1 433.
[10] 邹媛媛,刘洋,赵亮,等.两种抗病性不同的水稻种子固有细菌群落的多样性研究[J].中国农业科技导报,2016,18(4):9-16.
[11] 符鲜,杨树青,刘德平,等.套作小麦/玉米不同施氮水平对土壤养分与微生物数量的影响[J].干旱区研究,2017,34(1):43-50.
[12] RANIA A B A, JABNOUN-KHIAREDDINE H, NEFZI A, et al. Biocontrol of fusarium wilt and growth promotion of tomato plants using endophytic bacteria isolated fromSolanumelaeagnifoliumstems[J]. Biological Control, 2016,97:80-88.
[13] 卢昕,黄贵修.植物内生细菌生防作用研究进展[J].热带生物学报,2007,13(4):28-33.
[14] CHEBOTAR V K, MALFANOVA N V, SHCHERBAKOV A V, et al. Endophytic bacteria in microbial preparations that improve plant development (review) [J]. Applied Biochemistry & Microbiology, 2015,51(6):283-289.
[15] 林占熺,林冬梅,孙红英,等.菌草与大豆混种降低大豆产量但提高系统经济效益——以中国援卢旺达农业技术示范项目为例[J].福建农林大学学报(自然科学版),2015,44(6):624-628.
[16] CHAUDHARY H J, PENG G X, HU M, et al. Genetic diversity of endophytic diazotrophs of the wild rice,Oryzaaltaand identification of the new diazotroph, Acinetobacteroryzaesp. nov[J]. Microbial Ecology, 2012,63(4):813-821.
[17] SARATHAMBAL C, ILAMURUGU K, BALACHANDAR D, et al. Characterization and crop production efficiency of diazotrophic isolates from the rhizosphere of semi-arid tropical grasses of India[J]. Applied Soil Ecology, 2015, 87:1-10.
[18] 李志文.巨菌草作为能源草的特性研究[J].农业工程技术,2013(6):12-15.
[19] 曹荣,刘淇,赵玲,等.基于高通量测序的牡蛎冷藏过程中微生物群落分析[J].农业工程学报,2016,32(20):275-280.
[20] 赵帅,周娜,赵振勇,等.基于高通量测序分析盐角草根部内生细菌多样性及动态规律[J].微生物学报,2016,56(6):1 000-1 008.
[21] 林兴生,林占熺,林冬梅,等.荒坡地种植巨菌草对土壤微生物群落功能多样性及土壤肥力的影响[J].生态学报,2014,34(15):4 304-4 312.
[22] 郑贺云,黎志坤,李超,等.新疆阿克苏地区盐碱地细菌类群多样性及优势菌群分析[J].微生物学通报,2012,39(7):1 031-1 043.
[23] 任丽娟, 何聃, 邢鹏,等. 湖泊水体细菌多样性及其生态功能研究进展[J].生物多样性,2013,21(4):421-432.
[24] 陈倩.根际细菌的固氮、抗病与促生潜能研究[D].北京:中国农业科学院,2011.
[25] 蒲强,谭志远,彭桂香,等.根瘤菌分类的新进展[J].微生物学通报,2016,43(3):619-633.
[26] 张翠芳, 潘存德, 王世伟,等. 内生固氮菌DENB20在不同荒漠植物种体内的定殖动态[J].中国农学通报, 2017,33(6):123-128
[27] 刘振东,王波,薛蓓,等.高原作物青稞根及根际内生细菌群落结构的高通量分析[J].作物杂志,2016(6):49-52.
