马尾松茎叶中矿质元素含量及相关酶活性对MeJA的响应
2018-06-06姚瑞玲李慧娟张烨张晓宁王胤
姚瑞玲,李慧娟,张烨,张晓宁,王胤
(广西林业科学研究院 广西优良用材林资源培育重点实验室,广西 南宁530002)
马尾松(PinusmassonianaLamb.)是我国南方主要的造林树种,在广西、福建、云南、贵州、四川等16个省区均有分布,其利用价值高、用途广泛,具有广阔的推广应用前景[1-2]。马尾松的虫害危害严重,发生面积最大,其在我国马尾松分布区会周期性地暴发成灾,制约了马尾松产业的高效发展[3]。众多学者对马尾松抗虫性方面开展了大量研究[4-7],但有关马尾松诱导抗性机制方面的研究仍少见。
茉利酸甲酯(MeJA)属茉莉酸类物质,存在于植物体中的一种生长调节剂[8]。MeJA对植物有明显的生理效应,作为内源信号分子不仅调节植物的生长发育,而且通过外源应用能够诱导植物对机械伤害、病虫害、干旱、盐渍和低温等逆境的防御反应[9-11]。矿质元素与植物的生长和生理代谢活动密切相关[12]。研究发现,萜烯化合物影响着马尾松的抗虫能力[4-5,7],在MeJA处理后,其植株中萜烯类物质含量增加,马尾松抗虫性增强[6,13],但MeJA对马尾松茎叶中矿质元素吸收和累积有何影响仍不明确。本研究以马尾松幼苗为研究对象,采用电感耦合等离子体原子发射光谱法(ICP-AES),测定并分析了MeJA处理下诱导的马尾松幼苗植株茎叶中矿质元素含量及相关酶活性的变化,旨在明确在MeJA影响下马尾松产生诱导抗性时其矿质元素的响应机制,以期为今后开展马尾松对虫害诱导抗性研究提供参考。
1 材料与方法
1.1 试验材料
选择长势良好、高度为20-30cm,未受虫害、无机械损伤的盆栽健康马尾松幼苗(种质资源来自广西宁明县派阳山林场桐棉松优良林分),喷洒茉莉酸甲酯(茉莉酸甲酯购买于Sigma-Aldrich Chem.Co.,纯度>95%)。
1.2 试验设计
2015年1月至12月,先后采用不同浓度MeJA喷洒马尾松植株。马尾松毛虫(DendrolimuspunctatusWalker)取食性及其相关内源次生代谢物质组分变化的观测分析预备试验见图1。
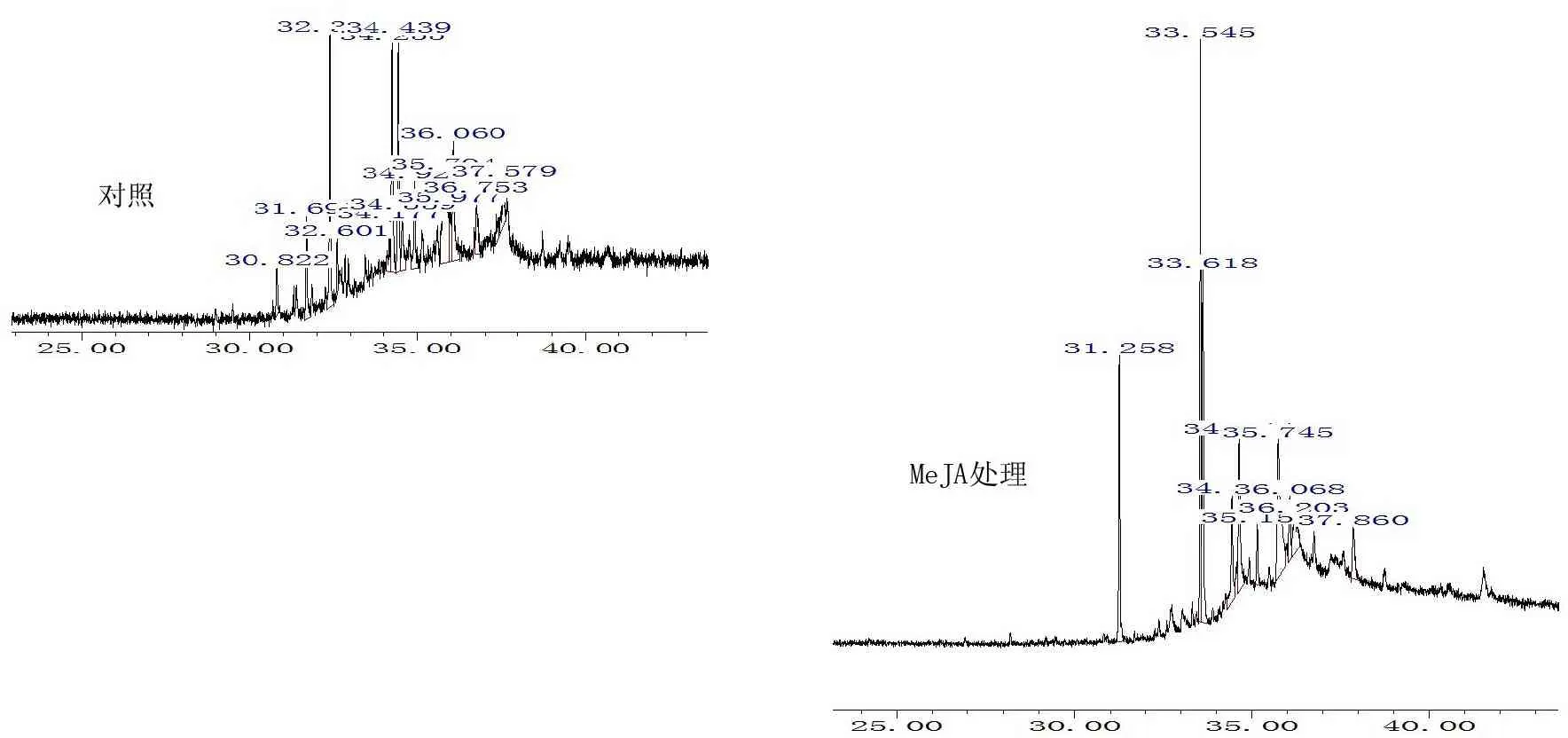
图10.2mmol/LMeJA处理4周马尾松幼苗针叶中萜烯类物质的总离子流气相色谱图
Fig.1 Total ionic chromatogram of terpenoids inPinusmassonianaseedling needles after four weeks of 0.2mmol/L MeJA treatment
根据预试验的分析结果,本研究选择处理后GC-MS分析萜类组分相对含量变化较大(图1),马尾松毛虫拒食性明显,且植株受伤害程度相对较轻的MeJA试验处理浓度,即以0.2mmol/L(pH7.0,含0.01%吐温-20)MeJA为处理、以蒸馏水(pH 7.0,含0.01%吐温-20)为对照(Control)。2016年4月至5月,将处理溶液叶面喷施马尾松幼苗,每次喷施溶液量100mL/盆(盆的直径×高为9cm×15cm),每个处理重复8盆,共处理16盆。连续处理幼苗4周,每2d喷施MeJA溶液1次,共喷施14次,停止喷施MeJA溶液5d后采样测定。采样时,对照、MeJA处理各选取8株幼苗,每个处理每个重复选取1株,每株选择处理进行期间植株上健康、完全伸展的成熟针叶束。本试验在人工气候箱(PQX-250)中进行,温度25℃/20℃(昼/夜),光周期12h,光强6 000Lx,相对湿度85%。
1.3 矿质元素含量测定
将马尾松幼苗整株植株洗净后,去掉根系再分成茎干及针叶两部分于80℃烘箱中烘干至恒重,将烘干材料研磨成碎末状,准确称取植株各部位的干粉末样品0.5g进行消煮,制成样液用于矿质元素的测定。矿质元素含量测定参照姚瑞玲等[14]的方法。硝态氮和铵态氮测定采用迟荪琳等[15]的方法。
1.4 酶活性测定
硝酸还原酶(NR)、亚硝酸还原酶(NiR)、谷氨酰胺合成酶(GS)和谷氨酸合成酶(GOGAT)4种酶活性测定参照张智猛等[16]的方法。
1.5 数据处理
采用SPSS 19.0统计分析软件进行数据分析及α=0.05和α=0.01水平方差分析及差异性显著检验。
2 结果与分析
2.1 矿质元素含量变化
从表1可以看出,MeJA处理后马尾松幼苗茎干中大量元素含量均无明显变化,但针叶中大量元素含量发生了改变。其中,大量元素中N、P、Mg的含量在MeJA处理后显著增加,尤其是N元素含量达到差异极显著水平,相较对照增加了109%。相对而言,K、Ca含量无明显变化。而经检测的B、Mn、Fe、Zn、Cu、Mo这6种微量元素,在MeJA处理前后其含量在马尾松幼苗茎干中变化不大,但针叶中Mn、Fe、Cu、Mo微量元素的含量在MeJA处理后显著增加,而B、Zn含量变化不明显。
从矿质元素在马尾松幼苗不同组织部位的变化来看,以针叶的变化最为明显。从马尾松茎叶中不同矿质元素的变化规律来看,茎干、针叶中N、P、Mg、Mn、Fe、Cu、Mo含量在MeJA处理下整体上均呈上升趋势,尤其是N含量变化极为显著。这说明,进行马尾松活体植株MeJA处理时,针叶中的大量元素含量变化相较茎干更为灵敏,并以N素的变化最大。

表1 MeJA处理下马尾松幼苗茎叶中营养元素含量变化
注:表中小、大写字母分别表示各矿质元素含量在α=0.05和0.01水平时同一植株组织中对照与处理间的差异显著性,下同。
2.2 N素代谢及相关酶活性变化
为进一步观测MeJA处理下马尾松植株茎叶中N元素含量变化情况,分别对其硝态氮、铵态氮的含量进行了分析,见表2。
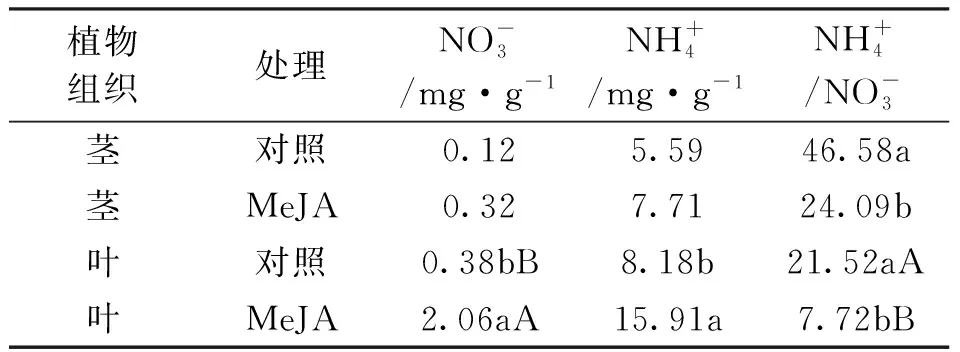
表2 MeJA处理下马尾松幼苗茎叶中硝态氮和铵态氮含量的变化
从表2中可知,与对照相比,供试幼苗茎干中硝态氮、铵态氮含量无明显变化,但铵硝比变化显著,在MeJA处理后,该比值明显下降。从针叶中N素代谢来看,硝态氮、铵态硝含量及铵硝比在MeJA处理下均变化显著,其中硝态氮含量和铵硝比达差异极显著水平。相较对照,针叶中硝态氮、铵态氮含量增加,铵硝比降低。
整体上,MeJA处理后马尾松幼苗茎干和针叶中硝态氮和铵态氮含量增加,而铵硝比降低,但其变化趋势以针叶部位较为明显。从N素含量变化而言,MeJA处理下硝态氮增加幅度较铵态氮大。
硝酸还原酶、亚硝酸还原酶、谷氨酰胺合成酶、谷氨酸合成酶是无机态氮转化为有机态氮的4种关键酶,直接影响植株中N素的代谢情况。由图2中MeJA处理后马尾松幼苗茎叶中4种N元素代谢酶活性变化来看,在MeJA诱导下,茎叶中硝酸、亚硝酸还原酶活性显著增加了,而谷氨酰胺、谷氨酸合成酶活性仅表现为在针叶中活性显著增加,在茎杆中变化不明显。
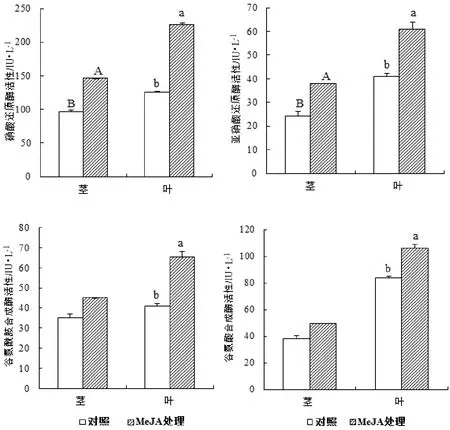
图2 MeJA处理下马尾松幼苗茎叶中N素代谢相关酶活性变化
3 结论与讨论
N、P、K、Ca、Mg、B、Mn、Fe、Zn、Cu、Mo是植物生长所必需的营养元素,它们通过直接参与植物体内重要有机化合物、植物激素、生物酶的合成或作为酶的活化剂,促进植物的光合作用及呼吸作用,氮代谢、碳水化合物代谢,调节渗透压及增加植物抗逆性等代谢活动影响植物的生根过程[12,17]。本试验研究发现,通过外源喷施MeJA 4周,马尾松幼苗植株茎叶中N、P、Mg、Mn、Fe、Cu、Mo等元素含量均呈上升趋势。N是植物生长发育所必需的重要元素,被称为“生命的元素”,是构成叶绿素、酶、ATP等化合物的元素;P是核酸的主要组成成分,是构成ATP的元素;Mg和Fe是合成叶绿素所必需的,主要影响植株的光合作用;Mn在光合作用中具有重要作用,植物缺锰,光合作用就明显降低;Cu是多种酶的重要组分,能通过调节多种酶的活性来影响植物生理代谢过程。Mo是固氮酶和硝酸还原酶的成分,氮代谢和豆科(Leguminosae)植物共生固氮与Mo密切相关[17-21]。从这几种矿质元素的生理功能来看,都与植物的光合作用生理代谢活动有关。以往研究证明,在MeJA诱导下,马尾松植株中诱导抗性增强,挥发性萜烯类物质增加[6-7]。马尾松产脂能力强,萜烯类化合物代谢旺盛,其中以挥发性单萜烯类物质含量最高[22]。从萜烯类物质的合成代谢途径来看,单萜烯类物质合成代谢场所主要为质体(包括叶绿体、白色体、有色体),合成前体为葡萄糖,与光合作用密切相关[23-27]。因此,在本试验研究中,相关矿质元素的含量变化是否是会改变马尾松植株的光合能力,从而改变萜烯类物质的合成代谢,进而实现马尾松诱导抗性能力增强,这需要进一步研究证实。
在养分代谢中,茎干主要负责植物养分的运输,而叶片则是植物合成与积累养分的重要部位[15]。在外源喷施MeJA处理下,本试验中马尾松幼苗植株茎干和针叶中矿质元素含量的变化以针叶较为显著。叶片是林木光合作用的主要器官,叶中营养元素含量最能反映植株新陈代谢的特点,最有代表性[28]。结合矿质元素变化结果来看,这说明今后在进行马尾松诱导抗性机制研究中,为科学、准确地反映试验研究结果,取样时应以针叶作为研究对象。
在本试验研究中,根据不同矿质元素含量对MeJA响应的变化趋势来看,以N素变化最为显著。从N素含量变化结果来看,在MeJA处理下硝态氮与铵态氮含量都增加了,但硝态氮含量的增幅明显比铵态氮含量的大。植株中N素代谢是从硝态氮铵态氮有机氮的过程,要实现无机氮到有机氮的转化,与硝酸还原酶、亚硝酸还原酶、谷氨酰胺合成酶和谷氨酸合成酶的活性密不可分[29-31]。通过外源喷施0.2mmol/L的MeJA溶液4周,马尾松幼苗植株茎叶中硝酸、亚硝酸还原酶活性明显增强,这可能是MeJA诱导下增加的硝态氮激发了硝酸和亚硝酸还原酶活性所致;而茎干中谷氨酰胺与谷氨酸合成酶活性偏低,则可能与茎干中铵态氮含量较低有关。这就是说,在MeJA处理下,马尾松幼苗植株中硝酸、亚硝酸还原酶与谷氨酰胺、谷氨酸合成酶的活性分别与硝态氮、铵态氮含量呈正相关的变化关系。在开展马尾松抗虫能力的调控中,适当增加植株中硝态氮、铵态氮含量,并以较低的能诱导N元素代谢酶活性增强,加快N元素转化与利用,有效发挥N素在马尾松诱导抗性中的作用。
参考文献:
[1] 丁贵杰.马尾松人工林生物量和生产力研究-I.不同造林密度生物量及密度效应[J].福建林学院学报,2003,23(1):34-38.
[2] 丁贵杰,周志春,王章荣,等.马尾松纸浆用材林培育与利用[M].北京:中国林业出版社,2006:1-10.
[3] 戈峰,李镇宇,谢映平,等.我国主要松树诱导抗虫性的一些规律比较[J].北京林业大学学报,2002,24(3):61-65.
[4] 胡少波,冯远斌,韦成礼,等.马尾松抗松毛虫植株的抗性研究[J].广西植保,1988(4):47-48.
[5] 马尾松抗松毛虫抗性研究组.马尾松抗松毛虫植株的抗性机制研究[J].林业科学,1990,26(2):133-141.
[6] 胡永建,任琴,李镇宇,等.虫害马尾松(PinusmassonianaLamb.)邻枝针叶挥发物及其内源茉莉酸甲酯的快速变化[J].生态学报,2008,28(11):5331-5337.
[7] 胡永建.虫害马尾松邻株信号通讯及茉莉酸甲酯诱导的蛋白表达差异研究[D].北京:北京林业大学,2010.
[8] Creelman R A,Mullet J E.Biosynthesis and action of jasmonate in plants[J].Annual Review Plant Physiology and Plant Molecular Biology,1997,48:355-381.
[9] 张秋菊,张爱华,孙晶波,等.植物体中萜类物质化感作用的研究进展[J].生态环境学报,2012,21(1):187-193.
[10] Ananieva K,Ananiew E D,Mishev K,etal.Methyl jas-monate is a more effective senescence-promoting factor inCucurbitapepo(zucchini ) cotyledons when compared with darkness at the early stage of senescence[J].Journal of Plant Physiology,2007,164:1179-1187.
[11] Hung K T,Kao C H.Nitric oxide acts as an antioxi-dant and delays methyl jasmonate-induced senes-cence of rice leaves[J].Journal of Plant Physiology,2004,161:43-52.
[12] 潘瑞炽.植物生理学(第七版)[M].北京:高等教育出版社,2012.
[13] 王立春,任琴,许志春,等.茉莉酸甲酯对马尾松松针萜烯类挥发物及马尾松毛虫生长发育的影响[J].北京林业大学学报,2008,30(1):79-84.
[14] 姚瑞玲,项东云,陈健波,等.2种桉树插穗生根过程中营养元素含量的变化[J].安徽农业科学,2010,38(33):18992-18993.
[16] 张智猛,万书波,宁堂原,等.氮素水平对花生氮素代谢及相关酶活性的影响[J].植物生态学报,2008,32(6):1407-1416.
[17] 刘忠新,刘莉梅.浅议植物生长所必须的营养元素与其生理功能[J].农村实用科技信息,2007(12):8.
[18] 宋纯鹏.植物衰老生物学[M].北京:北京大学出版社,1998.
[19] 曹仪植,宋占午.植物生理学[M].兰州:兰州大学出版社,1998.
[20] 李俊英.铜锌元素及酶活性与华北落叶松抗逆性的研究[D].太谷:山西农业大学,2003:63-66.
[21] 武维华.植物生理学[M].北京:科学出版社,2003.
[22] 安宁,丁贵杰.广西马尾松松脂的化学组成研究[J].中南林业科技大学学报,2012,32(3):59-62.
[23] Arigoni D,Sagner S,Latzel C,etal.Terpenoid biosynthesis from 1-deoxy-D-xylulose in higher plants by intramolecular skeletal rearrangement[C].Proceedings of the National Academy of Science of USA,1997,94:10600-10605.
[24] Bartram S,Jux A,Gleixner G,etal.Dynamic pathway allocation in early terpenoid biosynthesis of stress-induced lima bean leaves[J].Phytochemistry,2006,67:1661-1672.
[25] Eisenreich W,Rohdich F,Bacher A.Deoxyxylulose phosphate pathway to terpenoids[J].Trends in Plant Science,2010,6:78-84.
[26] 龚治,李典谟,张真.针叶树萜类合成酶研究进展[J].林业科学,2010,46(1):123-130.
[27] 王凌健,方欣,杨长青,等.植物萜类次生代谢及其调控[J].中国科学-生命科学,2013,43(12):1030-1046.
[28] 侯学煜.中国植被地理及优势植物化学成分[M].北京:科学出版社,1982.
[29] 武丽,张西仲,唐兴贵,等.钼胁迫对烟草含钼酶和碳氮代谢关键酶的影响[J].核农科学,2015,29(12):2385-2393.

[31] 徐春梅,陈丽萍,王丹英,等.低氧胁迫对水稻幼苗根系功能和氮代谢相关酶活性的影响[J].中国农业科学,2016,49(8):1625-1634.
