濒危植物杜鹃红山茶种群结构和动态变化
2018-06-06李辛雷孙振元李纪元殷恒福范正琪
李辛雷,孙振元,李纪元,殷恒福,范正琪,徐 翊,罗 建
(1. 中国林业科学研究院亚热带林业研究所,浙江 杭州 311400;2. 中国林业科学研究院林业研究所 林木遗传育种国家重点实验室,北京 100091;3. 广东阳春鹅凰嶂省级自然保护区,广东 阳春 529600)
杜鹃红山茶(CamelliaazaleaWei)隶属于山茶科(Theaceae)山茶属(CamelliaLinn.)[1],又名杜鹃叶山茶,仅分布于广东阳春鹅凰嶂省级自然保护区内,分布范围小于100 km2,已被《中国物种红色名录》[2]列为极危物种。杜鹃红山茶花期为4月份至12月份,但在适宜的栽培条件下一年四季均可开花,具有重要的观赏价值,是培育四季茶花的优良亲本[3-4]。杜鹃红山茶分布范围狭窄且生境恶劣,加之人为无序采挖,天然种群个体数量仅存1 000余株,处于极度濒危状态。
种群结构反映了种群数量动态、发展趋势及其与环境的相互关系[5-6]。研究濒危植物的种群结构及其动态变化有利于濒危植物保护策略制定,进而达到保护其自然资源的目的[7-8]。目前,关于山茶属植物种群生态学的研究主要集中在种群结构及空间分布格局[9-12]等方面,而对杜鹃红山茶种群结构动态的变化规律还有待深入研究。鉴于此,作者以杜鹃红山茶天然种群为研究对象,分析其种群结构和静态生命表,并基于静态生命表绘制该种群的个体标准化存活曲线、个体死亡率曲线和个体损失度曲线,对该种群的时间序列进行预测,探讨杜鹃红山茶种群动态变化及发展趋势,以期为杜鹃红山茶天然种群的保护及适宜生境的恢复提供基础研究资料。
1 研究区概况和研究方法
1.1 研究区概况
杜鹃红山茶仅分布于广东阳春鹅凰嶂省级自然保护区,地理坐标为北纬21°50′36″~21°58′40″,东经111°21′29″~111°36′03″。该区域属南亚热带气候,年均温22.1 ℃,年降水量3 428.9 mm,空气相对湿度80%以上,年日照时数1 743.4 h,无霜期350 d。土壤为红黄壤,含沙量较高,pH 5.2~pH 6.0。
杜鹃红山茶沿红花潭河流两岸呈狭窄的带状分布,种群个体数量少,生境片断化严重,种群具较强的次生性质;杜鹃红山茶的主要伴生种类有铁榄〔Sinosideroxylonpedunculatum(Hemsl.) H. Chuang〕和大头茶〔Gordoniaaxillaris(Roxb.) Dietr.〕等,群落概况及生境条件详见文献[3]。
1.2 研究方法
1.2.1 样地划分和调查方法 分别于2009年至2012年的不同季节对杜鹃红山茶种群进行了4次野外调查,全面清点保护区内杜鹃红山茶的个体数量。在全面调查的基础上,根据种群分布范围及生境片断化状况,沿河岸设置9个面积100 m×5 m的代表性样地;采用相邻格子法,将各样地划分为面积5 m×5 m的小样方,对杜鹃红山茶进行每木调查,并用数显游标卡尺(精度0.01 mm,上海恒量量具有限公司)测量其基径(BD)。
1.2.2 种群结构特征分析 采用径级代替龄级的方法,分析杜鹃红山茶的种群结构特征。因杜鹃红山茶的主干不明显,根据其生物学特性及其他灌木种群的研究结果[13-14],以其基径划分径级[3]。以基径5 mm为径阶距,划分15个径级:Ⅰ级(BD<5 mm)、Ⅱ级(5 mm≤BD<10 mm)、Ⅲ级(10 mm≤BD<15 mm)、Ⅳ级(15 mm≤BD<20 mm)、Ⅴ级(20 mm≤BD<25 mm)、Ⅵ级(25 mm≤BD<30 mm)、Ⅶ级(30 mm≤BD<35 mm)、Ⅷ级(35 mm≤BD<40 mm)、Ⅸ级(40 mm≤BD<45 mm)、Ⅹ级(45 mm≤BD<50 mm)、Ⅺ级(50 mm≤BD<55 mm)、Ⅻ级(55 mm≤BD<60 mm)、ⅩⅢ级(60 mm≤BD<65 mm)、ⅩⅣ级(65 mm≤BD<70 mm)和ⅩⅤ级(BD≥70 mm)。统计各径级个体存活数,并根据公式“个体百分率=(各径级个体存活数/个体总存活数)×100%”计算各径级的个体百分率;并以径级为横坐标、个体百分率为纵坐标,绘制杜鹃红山茶种群径级结构图。
1.2.3 静态生命表编制及曲线绘制 参考甄江红等[14]的方法编制杜鹃红山茶种群静态生命表。以径级(基径)代替龄级,将径级按从小至大的顺序视为时间顺序,对所有样地内杜鹃红山茶的存活个体分级结果进行统计并将数据标准化。以静态生命表为基础,以径级为横坐标,个体标准化存活数、个体死亡率和个体损失度为纵坐标分别绘制个体标准化存活曲线、个体死亡率曲线和个体损失度曲线[15-17]。
1.2.4 种群数量动态的时间序列预测 参考种群数量动态的时间序列预测方法[16,18],对未来2、5和10年后杜鹃红山茶的种群数量进行预测。
1.3 数据处理和分析
采用EXCEL 2007软件进行相关数据的处理与分析。
2 结果和分析
2.1 杜鹃红山茶种群的径级结构分析
以广东阳春鹅凰嶂省级自然保护区杜鹃红山茶种群径级为横坐标、个体百分率为纵坐标绘制该种群的径级结构图,结果见图1。由图1可以看出:杜鹃红山茶种群的径级结构总体表现为中间高、两端低的分布状态,其中,Ⅳ级(15 mm≤BD<20 mm)至Ⅵ级(25 mm≤BD<30 mm)的个体存活数最多,此后随径级增大而逐渐减少。Ⅲ级(10 mm≤BD<15 mm)至Ⅶ级(30 mm≤BD<35 mm)的个体存活数占个体总存活数的75.2%,其中,Ⅳ级至Ⅵ级的个体存活数占个体总存活数的52.0%,而Ⅰ级(BD<5 mm)和Ⅱ级(5 mm≤BD<10 mm)的个体存活数仅占个体总存活数的3.3%,表现出小径级和大径级的个体存活数较少而中间径级的个体存活数较多的特征。由此可见,杜鹃红山茶种群主要以中间径级个体占优势,严重缺乏幼苗,呈衰退趋势,属于下降型种群。

Ⅰ:BD<5 mm;Ⅱ:5 mm≤BD<10 mm;Ⅲ:10 mm≤BD<15 mm;Ⅳ:15 mm≤BD<20 mm;Ⅴ:20 mm≤BD<25 mm;Ⅵ:25 mm≤BD<30 mm;Ⅶ:30 mm≤BD<35 mm;Ⅷ:35 mm≤BD<40 mm;Ⅸ:40 mm≤BD<45 mm;Ⅹ:45 mm≤BD<50 mm;Ⅺ:50 mm≤BD<55 mm;Ⅻ:55 mm≤BD<60 mm;ⅩⅢ:60 mm≤BD<65 mm;ⅩⅣ:65 mm≤BD<70 mm;ⅩⅤ:BD≥70 mm. BD:基径 Basal diameter.图1 广东阳春鹅凰嶂省级自然保护区杜鹃红山茶种群的径级结构Fig. 1 Structure of diameter class of Camellia azalea Wei population in Ehuangzhang Provincial Nature Reserve in Yangchun of Guangdong Province
2.2 杜鹃红山茶种群的静态生命表及生命过程分析
2.2.1 种群静态生命表 广东阳春鹅凰嶂省级自然保护区杜鹃红山茶种群的静态生命表见表1。由表1可以看出:在杜鹃红山茶种群中,Ⅲ级(10 mm≤BD<15 mm)至Ⅸ级(40 mm≤BD<45 mm)(中间径级)的个体存活数较多,Ⅰ级(BD<5 mm)和Ⅱ级(5 mm≤BD<10 mm)(小径级)以及Ⅻ级(55 mm≤BD<60 mm)至ⅩⅤ级(BD≥70 mm)(大径级)的个体存活数均较少,呈衰退趋势;个体存活数和个体标准化存活数在Ⅰ级和Ⅱ级较少,并随径级增大而增加,Ⅳ级(15 mm≤BD<20 mm)至Ⅵ级(25 mm≤BD<30 mm)个体标准化存活数最多;此后随径级增大而减少,与种群径级结构的变化趋势基本一致。该种群Ⅰ级至Ⅲ级的个体标准化死亡数、个体死亡率和个体损失度均为负值,Ⅳ级后呈波动增加的趋势,Ⅸ级(40 mm≤BD<45 mm)后随径级增大呈波动减少的趋势,老龄个体因接近其寿命极限而增多直至个体死亡。该种群个体区间寿命从Ⅰ级开始增加,在Ⅳ级和Ⅴ级(20 mm≤BD<25 mm)达到峰值,此后随径级增大而减少;个体总寿命在Ⅰ级最高,此后随径级增大而减少。个体期望寿命和个体存活率总体上随径级增大而减少,但由于Ⅰ级、Ⅱ级、Ⅷ级(35 mm≤BD<40 mm)和ⅩⅣ级(65 mm≤BD<70 mm)的个体存活数极少,导致对幼苗和老树的个体期望寿命和个体存活率估计过高。
表1广东阳春鹅凰嶂省级自然保护区杜鹃红山茶种群的静态生命表1)
Table1StaticlifetableofCamelliaazaleaWeipopulationinEhuangzhangProvincialNatureReserveinYangchunofGuangdongProvince1)

径级2) Diameter class2)axlxdxqxKxLxTxexSxⅠ(BD<5 mm)236-109-3.0-1.4915 539153.74.0Ⅱ(5 mm≤BD<10 mm)8145-436-3.0-1.43635 44937.64.0Ⅲ(10 mm≤BD<15 mm)32581-419-0.7-0.67915 0868.81.7Ⅳ(15 mm≤BD<20 mm)551 0001100.10.19454 2954.30.9Ⅴ(20 mm≤BD<25 mm)49890-110-0.1-0.19453 3503.81.1Ⅵ(25 mm≤BD<30 mm)551 0002910.30.48552 4052.40.7Ⅶ(30 mm≤BD<35 mm)397093640.50.75271 5512.20.5Ⅷ(35 mm≤BD<40 mm)19345-18-0.1-0.13541 0243.01.1Ⅸ(40 mm≤BD<45 mm)203632180.60.92546701.80.4Ⅹ(45 mm≤BD<50 mm)814500.00.01454162.91.0Ⅺ(50 mm≤BD<55 mm)8145550.40.51182711.90.6Ⅻ(55 mm≤BD<60 mm)590540.60.9631531.70.4ⅩⅢ(60 mm≤BD<65 mm)236180.50.727902.50.5ⅩⅣ(65 mm≤BD<70 mm)118-36-2.0-1.136633.53.0ⅩⅤ(BD≥70 mm)354541.04.027270.50.0
1)ax:x径级的个体存活数Survival number of individual inxdiameter class;lx:x径级开始时的个体标准化存活数Standardized survival number of individual at the beginning ofxdiameter class;dx:从x径级到x+1径级的个体标准化死亡数 Standardized death number of individual fromxdiameter class tox+1 diameter class;qx:从x径级到x+1径级的个体死亡率 Mortality rate of individual fromxdiameter class tox+1 diameter class;Kx:x径级的个体损失度 Loss degree of individual inxdiameter class;Lx:从x径级到x+1径级的个体区间寿命 Span life of individual fromxdiameter class tox+1 diameter class;Tx:x径级及以上各径级的个体总寿命 Total life of individual fromxdiameter class to higher diameter class;ex:x径级的个体期望寿命Life expectancy of individual inxdiameter class;Sx:x径级的个体存活率 Survival rate of individual inxdiameter class.
2)BD:基径 Basal diameter.
2.2.2 生命过程分析 以静态生命表为基础,绘制该种群的个体标准化存活曲线、个体死亡率曲线和个体损失度曲线,结果见图2。结果(图2-A)显示:依据Deevey[19]对种群存活曲线类型的划分标准,杜鹃红山茶种群的个体标准化存活曲线为凸型曲线,属于Deevey Ⅰ型,其Ⅰ级和Ⅱ级的个体标准化存活数较少,Ⅱ级以上的个体标准化存活数迅速增加;Ⅳ级和Ⅵ级的个体标准化存活数最高,其次为Ⅴ级,此后随径级增大迅速减少,个体死亡数逐渐增加;Ⅸ级后个体标准化存活数减少,个体死亡数增加的趋势均变缓,越接近生理寿命,个体标准化存活数越少,个体死亡数越高。
结果(图2-B,C)还显示:杜鹃红山茶种群的个体死亡率曲线和个体损失度曲线的变化趋势基本一致,Ⅰ级和Ⅱ级的个体死亡率和个体损失度基本不变,从Ⅲ级开始个体死亡率和个体损失度均缓慢增加,此后个体死亡率和个体损失度升高的趋势增强,但存在较为明显的波动,且无明显峰值;从Ⅻ级后,个体死亡率和个体损失度升高的趋势变缓直至降低;至ⅩⅤ级时,种群个体接近其生理寿命,出现死亡高峰。

Ⅰ:BD<5 mm;Ⅱ:5 mm≤BD<10 mm;Ⅲ:10 mm≤BD<15 mm;Ⅳ:15 mm≤BD<20 mm;Ⅴ:20 mm≤BD<25 mm;Ⅵ:25 mm≤BD<30 mm;Ⅶ:30 mm≤BD<35 mm;Ⅷ:35 mm≤BD<40 mm;Ⅸ:40 mm≤BD<45 mm;Ⅹ:45 mm≤BD<50 mm;Ⅺ:50 mm≤BD<55 mm;Ⅻ:55 mm≤BD<60 mm;ⅩⅢ:60 mm≤BD<65 mm;ⅩⅣ:65 mm≤BD<70 mm;ⅩⅤ:BD≥70 mm. BD:基径 Basal diameter.图2 广东阳春鹅凰嶂省级自然保护区杜鹃红山茶种群的个体标准化存活曲线(A)、个体死亡率曲线(B)和个体损失度曲线(C)Fig. 2 Standardized survival curve (A),mortality rate curve (B),and loss degree curve(C) of individual in Camellia azalea Wei population in Ehuangzhang Provincial Nature Reserve in Yangchun of Guangdong Province
2.3 杜鹃红山茶种群的时间序列预测
广东阳春鹅凰嶂省级自然保护区杜鹃红山茶种群的时间序列预测结果见表2。由表2可以看出:与该种群现存的个体存活数相比,在未来2和5年后,Ⅷ级(35 mm≤BD<40 mm)至ⅩⅣ级(65 mm≤BD<70 mm)的个体存活数均呈现增加的趋势,但ⅩⅤ级(BD≥70 mm)的个体存活数基本上无明显变化。在未来10年后,该种群个体将出现老龄化的趋势,Ⅹ级(45 mm≤BD<50 mm)至ⅩⅤ级的个体存活数增加,且径级越大增加趋势越明显。由此可见,杜鹃红山茶种群呈衰退趋势,随时间推移,老龄个体存活数将逐渐增加,但由于幼苗严重缺乏,幼龄个体相对较少,最终也将导致老龄个体急剧减少。
表2广东阳春鹅凰嶂省级自然保护区杜鹃红山茶种群的时间序列预测结果
Table2ResultoftimesequencepredictiononCamelliaazaleaWeipopulationinEhuangzhangProvincialNatureReserveinYangchunofGuangdongProvince
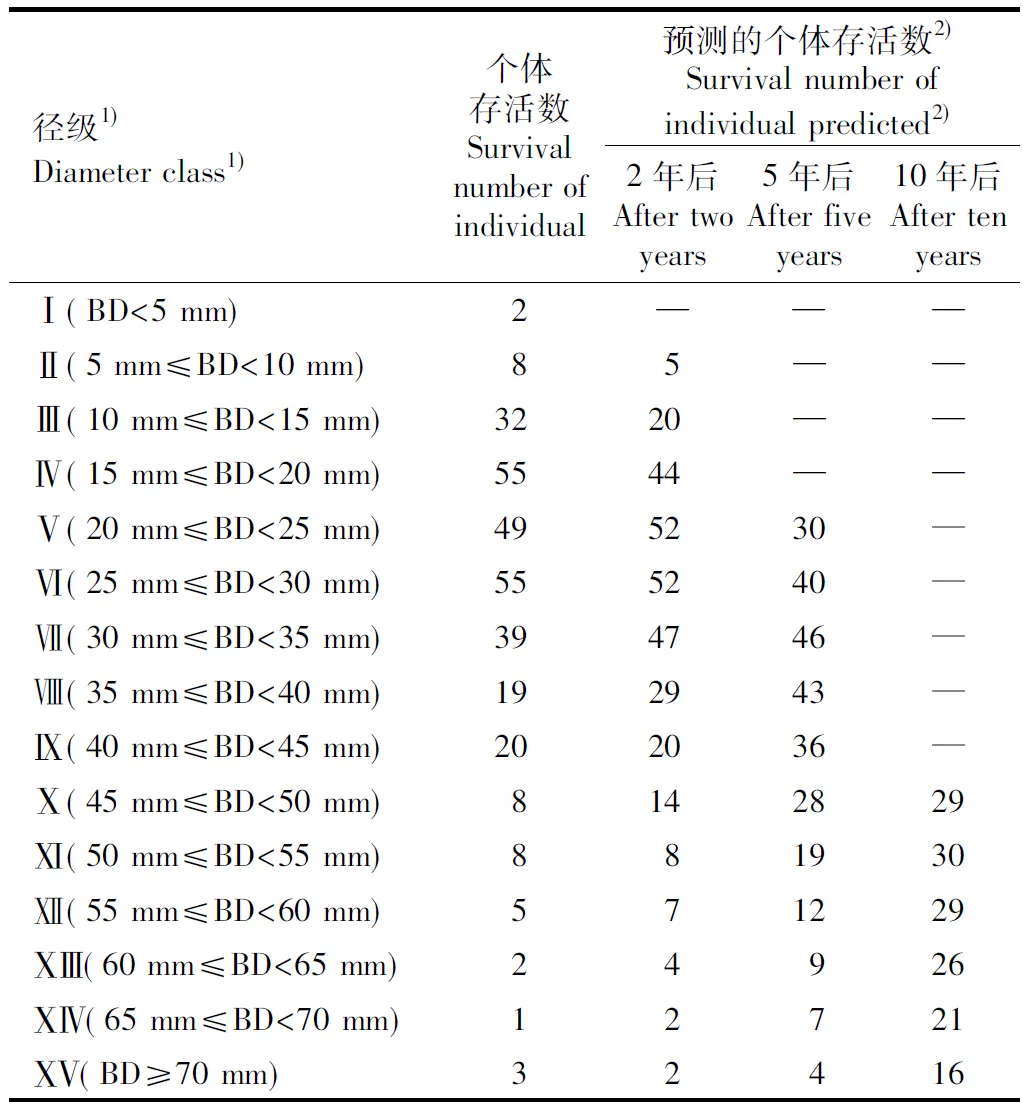
径级1)Diameter class1)个体存活数Survival number of individual预测的个体存活数2)Survival number of individual predicted2)2年后After two years5年后After five years10年后After ten yearsⅠ(BD<5 mm)2———Ⅱ(5 mm≤BD<10 mm)85——Ⅲ(10 mm≤BD<15 mm)3220——Ⅳ(15 mm≤BD<20 mm)5544——Ⅴ(20 mm≤BD<25 mm)495230—Ⅵ(25 mm≤BD<30 mm)555240—Ⅶ(30 mm≤BD<35 mm)394746—Ⅷ(35 mm≤BD<40 mm)192943—Ⅸ(40 mm≤BD<45 mm)202036—Ⅹ(45 mm≤BD<50 mm)8142829Ⅺ(50 mm≤BD<55 mm)881930Ⅻ(55 mm≤BD<60 mm)571229ⅩⅢ(60 mm≤BD<65 mm)24926ⅩⅣ(65 mm≤BD<70 mm)12721ⅩⅤ(BD≥70 mm)32416
1)BD:基径 Basal diameter.
2)—:无对应理论值 Without corresponding theoretical value.
3 讨论和结论
已有研究结果表明:种群静态生命表及以此为基础绘制的个体标准化存活曲线、个体死亡率曲线和个体损失度曲线整体上能够反映种群的数量动态变化趋势及结构特征[20-21]。通常种群径级结构呈纺锤型,存活曲线为凸型曲线,均表明种群趋向衰退[22]。本研究表明:杜鹃红山茶种群的Ⅲ级(10 mm≤BD<15 mm)至Ⅶ级(30 mm≤BD<35 mm)的个体存活数占个体总存活数的75.2%,而Ⅰ级(BD<5 mm)和Ⅱ级(5 mm≤BD<10 mm)的个体存活数仅占个体总存活数的3.3%,种群径级结构呈纺锤型;在种群静态生命表中,Ⅲ级至Ⅸ级(40 mm≤BD<45 mm)(中间径级)的个体存活数较多,Ⅰ级和Ⅱ级(小径级)及Ⅻ级(55 mm≤BD<60 mm)至ⅩⅤ级(BD≥70 mm)(大径级)的个体存活数较少,呈衰退趋势。由此可知,广东阳春鹅凰嶂省级自然保护区的杜鹃红山茶种群多为中间径级的个体,幼苗严重缺乏,为衰退型种群。Leak[23]指出,种群的存活曲线若为直线则表示该种群为稳定型,若为凹型则表示该种群为增长型,若为凸型则表示该种群为下降型。杜鹃红山茶种群的个体标准化存活曲线为凸型曲线,属于Deevey Ⅰ型[19]曲线,表明该种群为下降型种群。
作为种群动态的重要特征,个体死亡率曲线反映了个体生存率和个体死亡率随年龄变化的状况[24]。小径级的个体死亡率为负值,说明种群Ⅰ级和Ⅱ级的幼苗严重缺乏,种群呈衰退趋势[22]。金雅琴等[25]的研究结果表明:在朴树(CeltissinensisPers.)种群中,由于Ⅰ级和Ⅱ级的幼苗和幼树数量偏少,导致种群Ⅰ级和Ⅱ级的个体死亡率为负值,部分龄级存在不同程度的个体缺失,种群整体呈衰退趋势。 McEvoy等[26]认为,人为干扰破坏会导致种群密度降低,实生苗缺乏。本研究中,杜鹃红山茶种群Ⅰ级至Ⅲ级的个体死亡数、个体死亡率和个体损失度均为负值,也表明该种群的幼苗数量严重不足,种群若要持续发展需要补充相应数量的幼苗;其他径级的个体死亡数、个体死亡率和个体损失度出现负值或0,主要原因为人为干扰(如胡乱砍伐和挖掘等)导致部分径级个体缺失;Ⅳ级(15 mm≤BD<20 mm)后种群的个体死亡数、个体死亡率和个体损失度波动增加,主要是由于幼苗和幼树成长为成年树的过程中对养分及生存空间的需求增加,自疏和他疏作用增强,导致个体死亡数及个体死亡率大大增加;Ⅸ级后种群中的个体均已成年,总体数量趋于稳定,个体死亡数、个体死亡率和个体损失度降低。时间序列预测表明:杜鹃红山茶种群Ⅷ级(35 mm≤BD<40 mm)至ⅩⅣ级(65 mm≤BD<70 mm)的个体存活数表现出增加趋势,主要与种群中Ⅲ级至Ⅶ级的个体存活数较多有关,使未来种群中的中老龄个体得到充分补充。
在自然条件下,在种子萌发并发育成幼苗这一过程中受到的阻碍是植物濒危的主要原因[27],保持种群稳定性的关键是为种子萌发和幼苗生长提供适宜条件[28],且幼苗的存活情况也是种群能否成功更新的关键因子之一[29]。在野外条件下,杜鹃红山茶种群的结实率较高,具有产生大量幼苗的潜力。但由于杜鹃红山茶天然种群仅分布于河流两岸距河床5~10 m的河岸处,河床切割深、坡度较大,部分河段坡度达90°,生境条件恶劣;而且,杜鹃红山茶果实成熟后自然开裂,除部分种子被雨水冲走外,大多数种子散落在石缝中,因霉烂、空瘪而难以萌发,只有那些散落在适宜环境中的少数种子才能顺利萌发并形成幼苗,导致杜鹃红山茶天然种群幼苗严重缺乏。Lockhart等[30]认为,从维持种群持续稳定的角度出发,应该对种群中的实生苗进行保护,以维持种群丰富的遗传多样性。人为干扰(如无序砍伐和采挖等)不但导致杜鹃红山茶种群适宜生境破碎化,也对其种子萌发和幼苗生长有明显影响,进而阻碍种群更新。
综上所述,从种群的径级结构、个体标准化存活曲线和时间序列的预测结果,均可得出杜鹃红山茶种群“呈衰退趋势,为下降型种群”的研究结论,因此,应加强杜鹃红山茶天然种群保护区的建设,严禁无序采挖等人为干扰活动,同时对其生境条件及生态系统加以维护,从而促进杜鹃红山茶种群的自然更新。此外,在就地保护的基础上,通过采种育苗、扦插繁殖或组培快繁等方法进行人工繁育,建立种质资源圃,实施迁地保护;并对人工栽培群体进行仿生栽培、回归自然驯化及回迁等,从而保护并逐渐恢复杜鹃红山茶种群数量及其自然更新能力。
参考文献:
[1] 卫兆芬. 中国山茶属一新种[J]. 植物研究,1986,6(4):141-144.
[2] 汪 松,解 炎. 中国物种红色名录:第一卷[M]. 北京:高等教育出版社,2004:362.
[3] 李辛雷,孙振元,李纪元,等. 广东鹅凰嶂自然保护区杜鹃红山茶种群结构与空间分布格局[J]. 应用生态学报,2013,24(8):2115-2121.
[4] 高继银,PARKS C R,杜跃强. 山茶属植物主要原种彩色图集[M]. 杭州:浙江科学技术出版社,2005:34-35.
[5] 李先琨,苏宗明,向悟生,等. 濒危植物元宝山冷杉种群结构与分布格局[J]. 生态学报,2002,22(12):2246-2253.
[7] WU X P,ZHENG Y,MA K P. Population distribution and dynamics ofQuercusliaotungensis,FraxinusrhynchophllaandAcermonoin Dongling Mountain,Beijing[J]. 植物学报,2002,44(2):212-223.
[8] 陈少瑜,付玉嫔,吴 涛,等. 濒危植物大果木莲种群格局及濒危原因分析[J]. 植物资源与环境学报,2012,21(2):102-106.
[9] 张治国,王仁卿. 中国分布北界的山茶(Camelliajaponica)种群大小结构和空间格局分析[J]. 植物生态学报,2000,24(1):118-122.
[10] 操国兴,钟章成,刘 芸,等. 缙云山川鄂连蕊茶种群空间分布格局研究[J]. 生物学杂志,2003,20(1):10-12.
[11] 戴 月,薛跃规. 濒危植物顶生金花茶的种群结构[J]. 生态学杂志,2008,27(1):1-7.
[12] 柴 勇,朱 华,孟广涛,等. 云南哀牢山古茶树群落优势树种的种群结构与分布格局[J]. 林业科学研究,2011,24(3):277-284.
[13] 何恒斌,张惠娟,贾桂霞. 磴口县沙冬青种群结构和空间分布格局的研究[J]. 林业科学,2006,42(10):13-18.
[14] 甄江红,刘果厚. 不同生境条件下四合木(TetraenamongolicaMaxim.)种群结构特征[J]. 生态学报,2008,28(4):1829-1841.
[15] 熊 能,金则新,顾婧婧,等. 千岛湖次生林优势种种群结构与分布格局[J]. 生态学杂志,2010,29(5):847-854.
[16] 申仕康,马海英,王跃华,等. 濒危植物猪血木(EuryodendronexcelsumH. T. Chang)自然种群结构及动态[J]. 生态学报,2008,28(5):2404-2412.
[17] 张文辉,卢彦昌,周建云,等. 巴山北坡不同干扰条件下栓皮栎种群结构与动态[J]. 林业科学,2008,44(7):11-16.
[18] 张文辉,王延平,康永祥,等. 濒危植物太白红杉种群年龄结构及其时间序列预测分析[J]. 生物多样性,2004,12(3):361-369.
[19] DEEVEY E S,Jr. Life tables for natural populations of animals[J]. The Quarterly Review of Biology,1947,22(4):283-314.
[20] 陈远征,马祥庆,冯丽贞,等. 濒危植物沉水樟的种群生命表和谱分析[J]. 生态学报,2006,26(12):4267-4272.
[21] 张桥英,罗 鹏,张运春,等. 白马雪山阴坡林线长苞冷杉(Abiesgeorgei) 种群结构特征[J]. 生态学报,2008,28(1):129-134.
[22] 熊 能,金则新,陈 琢. 千岛湖姥山马尾松种群结构和分布格局研究[J]. 植物研究,2010,30(5):537-542.
[23] LEAK W B. An expression of diameter distribution for unbalanced uneven-aged stands and forests[J]. Forest Science,1964,10(1):39-50.
[24] 林 玥,任坚毅,岳 明. 太白山红桦种群结构与空间分析[J]. 植物生态学报,2008,32(6):1335-1345.
[25] 金雅琴,李冬林,孙晓龙,等. 南京方山朴树种群结构与分布格局研究[J]. 植物研究,2011,31(5):603-609.
[26] MCEVOY P M,MCADAM J H,MOSQUERA-LOSADA M R,et al. Tree regeneration and sapling damage of pedunculate oakQuereusroburin a grazed forest in Galicia,NW Spain:a comparison of continuous and rotational grazing systems[J]. Agroforestry System,2006,66(2):85-92.
[27] 谢宗强,陈伟烈,路 鹏,等. 濒危植物银杉的种群统计与年龄结构[J]. 生态学报,1999,19(4):523-528.
[28] 张文辉,祖元刚,刘国彬. 十种濒危植物的种群生态学特征及致危因素分析[J]. 生态学报,2002,22(9):1512-1520.
[29] 韩文娟,袁晓青,张文辉. 油松人工林林窗对幼苗天然更新的影响[J]. 应用生态学报,2012,23(11):2940-2948.
[30] LOCKHART B R,CHAMBERS J L. Cherrybark oak stump sprout survival and development five years following plantation thinning in the lower Mississippi alluvial valley,USA[J]. New Forests,2007,33(2):183-192.
