PSGL-1通过炎症反应在小鼠盐敏感性高血压中的作用机制研究
2018-05-31毛红亚姜晓亮杨志伟中国医学科学院医学实验动物研究所北京协和医学院比较医学中心北京100021
付 慧,毛红亚,姜晓亮,刘 星,刘 雪,杨志伟(中国医学科学院医学实验动物研究所,北京协和医学院比较医学中心,北京 100021)
高血压是一种常见的机体病症,威胁到世界20%~30%的人群健康,也是脑卒中、冠心病、心功能不全、肾病等人类重大疾病的主要危险因素之一。在原发性高血压群体中盐敏感者占50%~60%[1 - 3]。研究表明,盐敏感性高血压病人并发心脑血管事件的风险远远高于盐耐受性高血压,盐敏感性高血压已成为威胁人类健康的重要病理因素之一[4]。盐敏感性高血压的病理机制比较复杂,一般认为高盐饮食导致动脉血管收缩及肾脏钠水代谢潴留是其主要发病机制[5 - 6],但是通过舒张血管和利尿等药物治疗,仍有相当比例的高血压患者的血压并没能得到有效控制[7]。即使没有导致血压的变化,高盐饮食仍然增加了心血管疾病发生的风险[4]。近年来,随着慢性炎症反应在盐敏感高血压及其它心血管疾病发生发展中作用及其机制研究的进展,炎症反应可能是高盐饮食导致血压升高及其靶器官损伤的重要作用机制[8]。
P-选择素糖蛋白配体-1(PSGL-1)是表达于白细胞表面的跨膜糖化蛋白,与白细胞活化密切相关。PSGL-1与选择素(selectin)分子的相互作用在白细胞粘附起始阶段起重要作用[9 - 12],PSGL-1可以明显提高血小板活化因子诱导的白细胞活化,与整合素等细胞因子共同作用促进白细胞粘附[13]。研究表明,PSGL-1参与调节T淋巴细胞分化过程,放大炎症信号,是调节T淋巴细胞参与免疫反应的重要靶点[14]。白细胞释放的TNF-α、IL-1等炎症因子可以诱导内皮细胞和血小板内储存的选择素移位到细胞表面,与白细胞表面的PSGL-1结合,进一步促进白细胞粘附和炎症反应的发生发展[15]。因此,PSGL-1在炎症反应发生过程中发挥重要作用,但是否参与盐敏感性高血压的病理发展过程有待进一步研究。
为了检测PSGL-1在盐敏感性高血压发生发展中的作用及其相关机制,本研究利用PSGL-1基因敲除小鼠并通过高盐喂养,检测小鼠血压变化及机体中炎症反应和肾脏靶器官损伤情况,以探索盐敏感性高血压及其靶器官损伤新的病理机制,为高血压及其靶器官损伤的防治提供参考。
1 材料和方法
1.1 实验动物
SPF级PSGL-1基因敲除(PSGL-1-/-)小鼠及其野生型(PSGL-1+/+)对照小鼠由美国俄克拉荷马大学健康科学中心夏利军教授捐赠,实验动物均在中国医学科学院医学实验动物研究所SPF级环境饲养[SYXK (京) 2015-0035],动物实验方案获实验动物使用与管理委员会(IACUC)的批准[ILAS-PG-2014-006]。将8周龄、体重约20 g的雄性PSGL-1-/-和PSGL-1+/+小鼠各12只,分别给予高盐(6% NaCl)和正常盐(0.4% NaCl)喂养3个月[16],然后,在麻醉(0.6%戊巴比妥)状态下利用颈总动脉插管法测量小鼠血压,牺牲小鼠,取小鼠皮肤、肾脏等组织,用于炎症细胞浸润和相关炎症因子表达,以及靶器官损伤的检测。
1.2 主要试剂与仪器
IL-6单克隆抗体购自Cell Signaling Technology(#12912);TNF-α单克隆抗体购自Santa Cruz(SC-52746);IL-1β多克隆抗体购自Santa Cruz(SC-7884);CD3单克隆抗体购自Santa Cruz(SC-20047);CD68单克隆抗体购自Novus(NBP1-74570SS);CTGF多克隆抗购自Proteintech(23936-1-AP);BCA蛋白浓度测定试剂盒(增强型)购自碧云天(P0010);兔超敏二步法试剂盒购自中杉金桥(PV9001);小鼠超敏二步法试剂盒购自中杉金桥(PV9002);DAB显色试剂盒(20 ×)购自中杉金桥(ZLI-9019);CD45-APC/Cy7小鼠抗体购自Biolegend(103116)、CD3-PE/Cy7小鼠抗体购自Biolegend(100320)。
多通道生理仪(BIOPAC Systems, Inc.);血压检测仪(BP-2000 Series II, Visitech Systems);电泳仪(北京六一仪器厂,DYY-C);电泳槽(上海天能,VE-180);电转槽(上海天能,VE-186);多功能酶标仪(Multiskan GO);化学发光凝胶成像仪(Tanon-5500);正向显微镜(Leica DM6000 B);低温高速离心机(Beckman);流式细胞仪(BD FACSCantoTM)。
1.3 实验方法
1.3.1 免疫组化方法检测组织的炎症细胞浸润
取小鼠皮肤组织常规固定,将石蜡包埋的切片(4 μm)二甲苯脱蜡后在不同浓度梯度酒精溶液中由高向低浸泡浸水,并使用0.3%过氧化氢阻断内源性过氧化物酶活性。在柠檬酸缓冲液(pH 6.0)中通过微波热诱导表位回收法揭开抗原。用羊血清阻断非特异性结合,然后用目标单克隆抗体杂交,4℃温育过夜。用超敏二步法试剂盒孵育后DAB显色试剂盒镜下显色,苏木素染色,镜下观察。
1.3.2 Western blot方法检测肾脏组织中的蛋白表达
提取新鲜小鼠肾脏全蛋白,用BCA法检测蛋白浓度。采用10%聚丙烯酰胺凝胶电泳,流转至硝酸纤维素膜,封闭液(0.1%缓血酸铵缓冲液配制5%脱脂奶粉)室温封闭2 h,加特异性抗体4℃孵育过夜,0.1%缓血酸铵缓冲液洗膜3次后加辣根过氧化物标记的IgG室温孵育1 h,洗膜3次后,化学发光凝胶成像仪自动曝光显影后使用Image J软件分析灰度,结果以目的蛋白/β-actin灰度值比值表示蛋白质的相对表达水平。
1.3.3 流式细胞术检测主动脉血管中炎症细胞浸润
为了进一步验证炎症细胞的浸润情况,本研究剥离小鼠胸主动脉和腹主动脉血管,将其剪碎消化至单个细胞悬液后,加入流式抗体CD45-APC/Cy7、CD3-PE/Cy7,充分混匀后室温避光孵育30 min,上机检测小鼠主动脉血管中浸润的白细胞和T淋巴细胞的百分率。
1.4 统计学方法
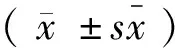
2 结果
2.1 PSGL-1参与高盐诱导的高血压发生发展
本研究给予PSGL-1-/-小鼠和PSGL-1+/+小鼠高盐(6% NaCl)或正常盐(0.4% NaCl)喂养,发现,对于PSGL-1+/+小鼠,高盐喂养组的收缩压、舒张压和平均脉压均明显高于正常盐喂养组(P< 0.05);而对于PSGL-1-/-小鼠,高盐喂养组和正常盐喂养组的血压并无明显变化(P> 0.05)(图1)。提示PSGL-1在盐敏感性高血压的发生发展过程中起重要作用。
2.2 PSGL-1参与高盐诱导的皮肤组织炎症细胞浸润的调节
本研究用免疫组织化学染色法对小鼠皮肤组织浸润T细胞和巨噬细胞特异性染色,发现在PSGL-1+/+小鼠高盐喂养后皮肤组织的T细胞和巨噬细胞浸润数量明显增多,而PSGL-1-/-小鼠高盐喂养后其T细胞和巨噬细胞浸润数量并未发现明显的变化(图2,图3)。表明PSGL-1在炎症细胞浸润过程中起重要作用,PSGL-1的缺失抑制高盐诱导的炎症细胞在组织中的浸润。

注:WT:PSGL-1+/+小鼠;KO:PSGL-1-/-小鼠;NS:0.4% NaCl;HS:6% NaCl。下同。与其它组小鼠比较,one-way ANOVA,* P< 0.05。图1 颈动脉插管法检测PSGL-1+/+小鼠和PSGL-1-/-小鼠动脉血压的变化Note.WT: PSGL-1+/+mice; KO: PSGL-1-/-mice; NS: 0.4% NaCl; HS: 6% NaCl. The same below. Compared with rats in the other groups, one-way ANOVA, * P< 0.05.Fig.1 Blood pressure in PSGL-1+/+ and PSGL-1-/- mice measured by carotid catheterization

注:标尺=50 μm。图2 T淋巴细胞在PSGL-1+/+和PSGL-1-/-小鼠皮肤组织的浸润情况Note.Bars=50 μm.Fig.2 Infiltration of T lymphocytes in the skin of PSGL-1+/+ and PSGL-1-/- mice
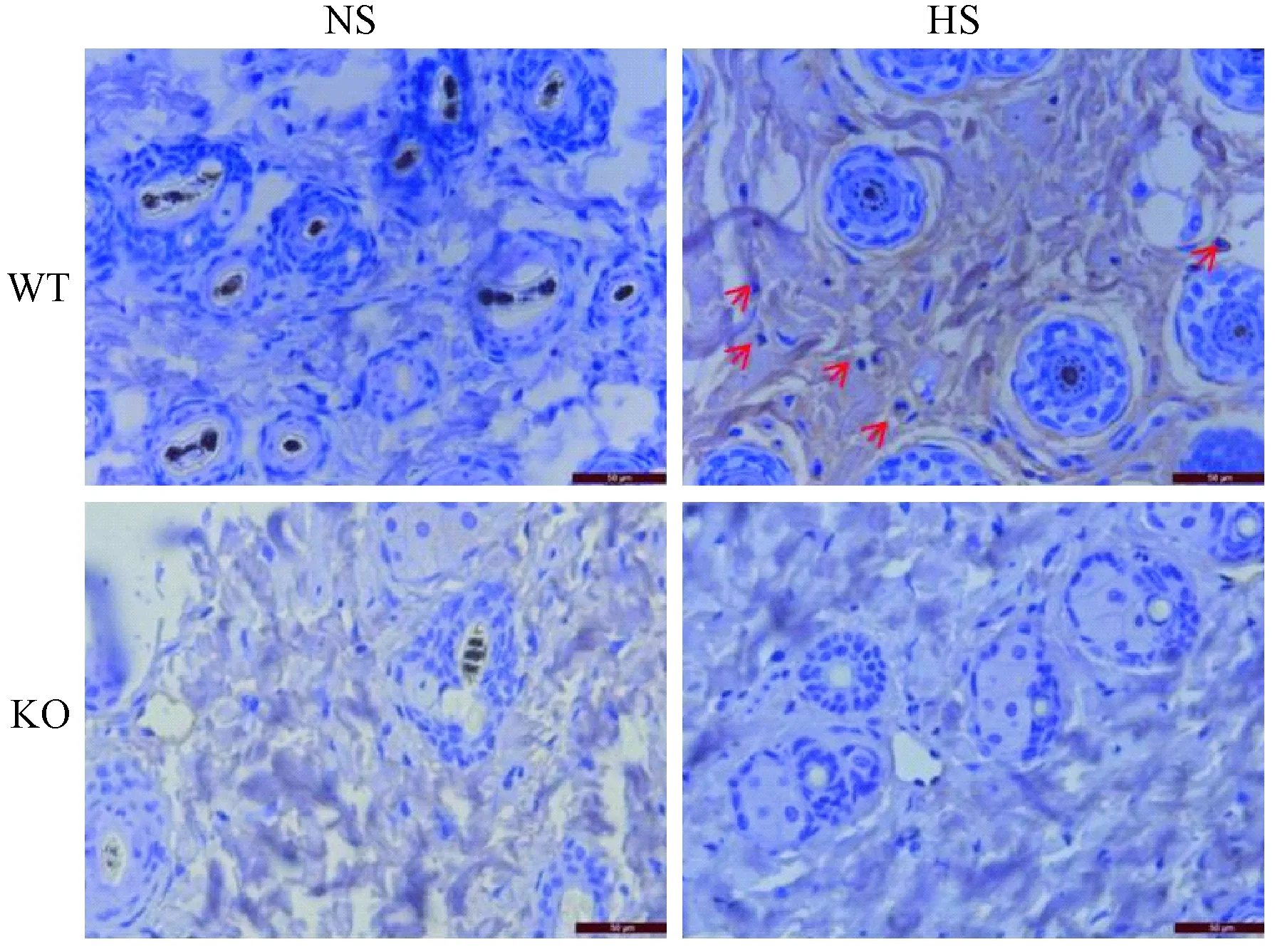
注:标尺=50 μm.图3 巨噬细胞在PSGL-1+/+和PSGL-1-/-小鼠皮肤组织的浸润情况Note.Bars=50 μm.Fig.3 Infiltration of macrophages in the skin of PSGL-1+/+ and PSGL-1-/- mice
2.3 PSGL-1参与高盐诱导的主动脉血管中炎症细胞浸润的调节
用流式细胞术检测小鼠主动脉血管中炎症细胞浸润数量,发现在PSGL-1+/+小鼠高盐喂养后小鼠主动脉血管中CD45+白细胞和CD3e+T细胞的浸润数量明显增多,而PSGL-1-/-小鼠高盐喂养后其浸润数量并没有出现明显变化(图4A,4B),证明PSGL-1参与高盐诱导的炎症细胞向主动脉中浸润的过程。PSGL-1缺失对此过程有抑制作用,从而抑制炎症反应的发生发展。
2.4 PSGL-1参与高盐诱导的肾脏炎症因子表达的调节
肾脏是机体血压调节的重要器官,为了进一步验证机体炎症反应发生情况,本研究检测了肾脏IL-6、IL-1β、TNF-α蛋白水平的表达。检测结果发现,PSGL-1+/+小鼠,高盐喂养组肾脏IL-6、IL-1β、TNF-α的表达明显高于正常盐喂养组;而PSGL-1-/-小鼠,高盐喂养组与正常盐喂养组肾脏IL-6、IL-1β、TNF-α表达无明显差异(图5A,5B,5C)。表明PSGL-1基因缺失对高盐诱导的肾脏炎症反应有抑制作用。
2.5 PSGL-1参与高盐诱导的肾脏纤维化水平的调节
为了检测高盐对肾脏损伤的影响,本研究检测了肾脏中作为肾脏损伤的标示蛋白结缔组织生长因子(connective tissue growth factor,CTGF)[17]的表达情况。检测发现,PSGL-1+/+小鼠高盐喂养组肾脏CTGF蛋白表达量明显高于正常盐喂养组;而PSGL-1-/-小鼠肾脏中,高盐喂养组CTGF蛋白的表达与正常盐喂养组无明显差异(图6)。结果表明,PSGL-1基因缺失对高盐诱导的肾脏纤维化具有抑制作用。
3 讨论
高盐摄入是盐敏感性高血压发生发展的重要因素,盐敏感性高血压发生发展的机理主要包括肾脏对钠水代谢调节、血管平滑肌的收缩、交感神经系统兴奋等[5, 18 - 19]。近年来,盐敏感性高血压被认为是一种低度炎症性疾病,免疫系统在盐敏感性高血压的发生发展中的作用引起研究者的重视。高盐等环境刺激下,作为与选择素(selectin)结合的配体,白细胞表面的P-选择素糖蛋白配体-1(PSGL-1)识别并结合表达在活化的内皮细胞表面的选择素,促进白细胞在血管内皮的粘附和浸润[9 - 10, 12, 20],促进炎症反应的发生发展。
本研究发现,通过高盐饮食诱发野生型小鼠发生盐敏感性高血压,且其麻醉状态下血压升高值与其它小鼠盐敏感高血压模型的血压升高值相近,如去氧皮质酮(deoxycorticosterone,DOCA)缓释诱发小鼠盐敏感性高血压模型[21],血管紧张素II诱导的高血压模型[22]等。人类收缩压低于120 mmHg时为理想血压,高于140 mmHg时为高血压[23],其差值与本实验中血压升高值相近。另外野生型小鼠高盐饮食后动脉血管和皮肤中炎症细胞(T细胞、巨噬细胞)浸润、炎症因子(IL-6、IL-1β、TNF-α)及肾脏纤维化标识蛋白(结缔组织生长因子,CTGF)表达增加,而PSGL-1敲除后小鼠并未发生以上现象。该结果提示PSGL-1可能通过炎症反应参与了盐敏感性高血压的发生发展及其靶器官的损伤。
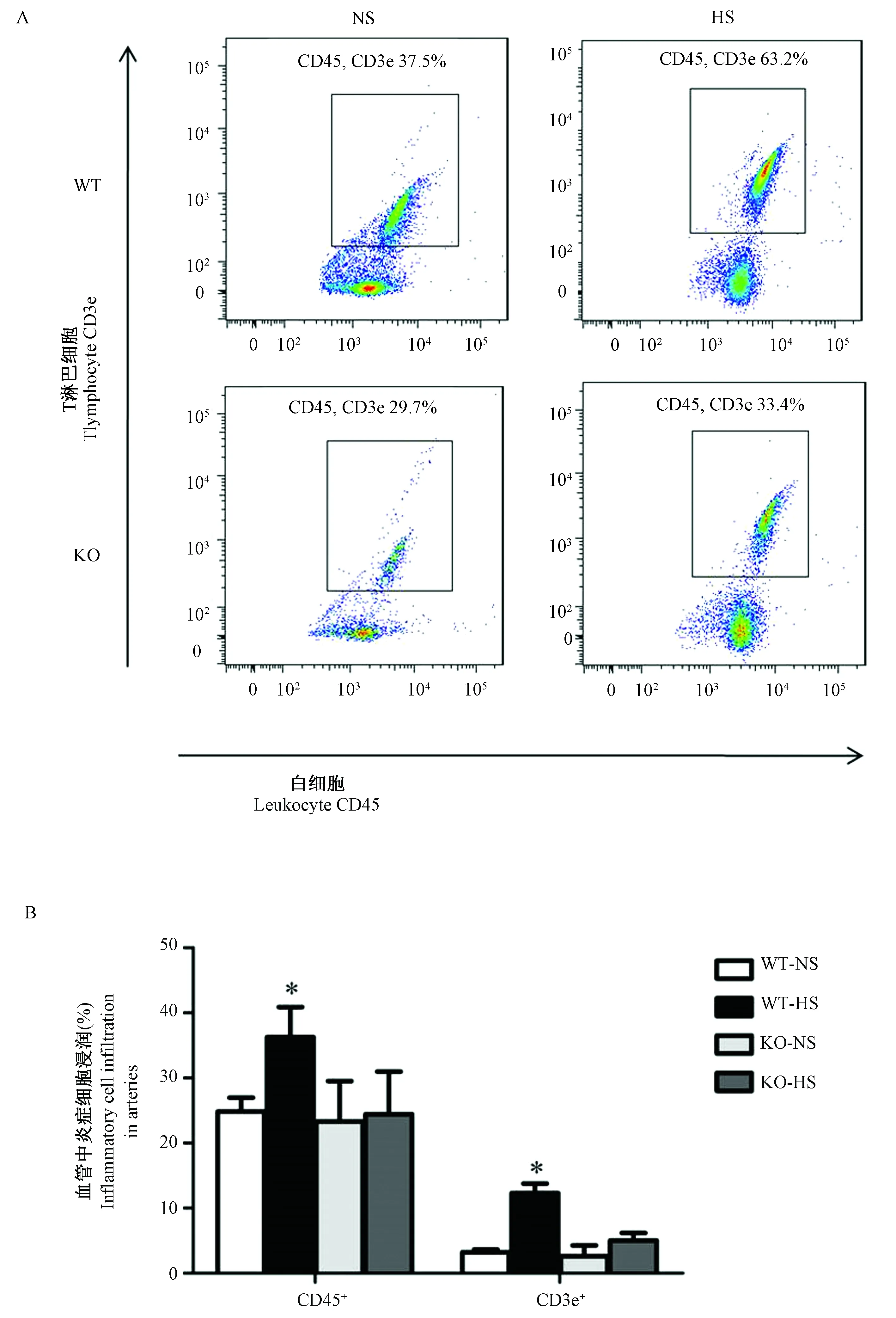
注:A:主动脉血管中浸润的炎症细胞图谱(流式细胞术散点图)。B:浸润的炎症细胞在主动脉血管细胞中的比例。与其它组小鼠比较,one-way ANOVA,* P< 0.05。图4 白细胞和T淋巴细胞在PSGL-1+/+和PSGL-1-/-小鼠主动脉血管中的浸润Note.A: Inflammatory cells in the aorta (flow cytometry scatter plot). B: Percentage of infiltrating inflammatory cells in aortic vascular cells. Compared with rats in the other groups, one-way ANOVA,* P< 0.05.Fig.4 Infiltration of leukocytes and T lymphocytes in the aorta of PSGL-1+/+ and PSGL-1-/- mice
有研究表明,高盐诱导Dahl盐敏感大鼠发生高血压同时,肾脏浸润性T淋巴细胞和巨噬细胞显著增加[11],肾脏中浸润的T淋巴细胞和巨噬细胞等炎症细胞分泌多种细胞因子[11, 24],直接或间接参与了盐敏感性高血压的发生发展和靶器官损伤[25 - 26]。IL-6、IL-1β和TNF-α均是炎症反应发生发展过程中发挥重要作用的炎症因子或促炎因子[27]。IL-6可通过进一步介导巨噬细胞在肾脏中的浸润增加或增殖[11, 28 - 29];IL-1β经炎症小体导致肾脏炎症和肾脏纤维化[30];IL-17激活RhoA/Rho激酶促进内皮功能障碍等参与高血压及靶器官损伤;TNF-α作用于血管内皮细胞促使其分泌其他炎症因子促进炎症反应[29],同时通过抑制eNOS表达以抑制血管舒张作用,或通过调节肾脏的血液动力学和排泄功能[31],促进高血压及靶器官损伤的发生发展[11, 32]。本研究发现,高盐喂养小鼠的血压明显升高,并伴随其皮肤T细胞、巨噬细胞浸润增多和肾脏组织中IL-6、IL-1β、TNF-α表达量升高,而PSGL-1缺陷小鼠高盐喂养后并未引起其血压、炎症细胞浸润和肾脏组织炎症因子表达量的明显变化,显示PSGL-1可能通过炎症反应参与盐敏感性高血压的发生发展,但具体的作用机制还有待于进一步研究。

注:A、B、C:分别为小鼠肾脏中IL-6、IL-1β、TGF-α蛋白的相对表达量。与其它组小鼠比较,one-way ANOVA,* P< 0.05。图5 PSGL-1+/+和PSGL-1-/-小鼠肾脏中IL-6、IL-1β和TNF-α蛋白表达Note.A, B, C: The expression of IL-6, IL-1β, TGF-α in the kidney of PSGL-1+/+ and PSGL-1-/- mice, respectively. Compared with rats in the other groups, one-way ANOVA, * P< 0.05.Fig.5 Interleukin-6, interleukin-1β, and tumor necrosis factor-α expression in the kidney of PSGL-1+/+ and PSGL-1-/- mice
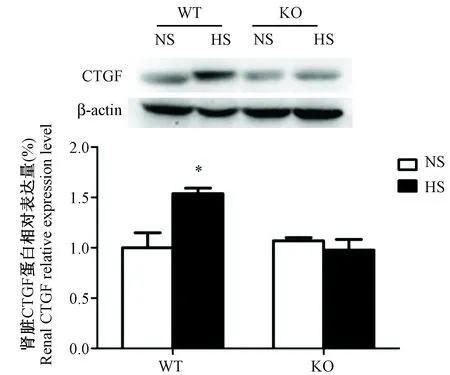
注:与其它组小鼠比较,one-way ANOVA,* P< 0.05。图6 PSGL-1+/+和PSGL-1-/-小鼠肾脏中CTGF蛋白的表达Note.Compared with rats in the other groups, one-way ANOVA,* P< 0.05.Fig.6 Connective tissue growth factor expression in the kidney of PSGL-1+/+ and PSGL-1-/- mice
结缔组织生长因子CTGF是一种炎症介质,CTGF的表达受炎症因子(如TNF-α,IL-1β,TGF-β等)及其它细胞外基质酶等多种炎症介质的调节[33],肾脏在发生损伤时CTGF表达升高。作为促纤维化因子TGF-β的下游信号介质,CTGF表达增加可能是器官纤维化发生发展的中心环节,CTGF与TGF-β结合增强TGF-β与其受体的二聚化[34],进一步促进TGF-β信号传导,促进组织纤维化,导致肾脏功能损伤[35]。CTGF能通过激活αvβ5整合素,凋亡信号调节激酶1,p38/JNK和AP-1/NF-κB通路,上调IL-6的表达,促进炎症反应[36]。本研究发现,在高盐喂养野生型小鼠发生高血压的同时,肾脏中CTGF的表达量明显增多,而PSGL-1缺失小鼠高盐喂养后血压和CTGF的表达量都无明显变化,表明PSGL-1缺失能够抑制高盐诱导的组织炎症反应发生而导致的血压升高和肾脏器官损伤。但是,高盐诱导的高血压和肾脏损伤之间的因果关系还有待进一步研究。
高血压控制的根本目的是靶器官损伤的预防与治疗。本研究揭示了PSGL-1通过调节炎症反应在高盐诱导的盐敏感性高血压的发生发展和靶器官损伤中的重要作用,并初步探索了PSGL-1通过调控T细胞和巨噬细胞浸润参与盐敏感性高血压的可能机制,为高血压的预防和治疗提供了新的理论依据。
参考文献:
[1] Franco V, Oparil S. Salt sensitivity, a determinant of blood pressure, cardiovascular disease and survival [J]. J Am Coll Nutr, 2006, 25(3 Suppl): 247S - 255S.
[2] Meneton P, Jeunemaitre X, de Wardener HE, et al. Links between dietary salt intake, renal salt handling, blood pressure, and cardiovascular diseases [J]. Physiol Rev, 2005, 85(2): 679 - 715.
[3] Weinberger MH, Fineberg NS, Fineberg SE, et al. Salt sensitivity, pulse pressure, and death in normal and hypertensive humans [J]. Hypertension, 2001, 37(2 Pt 2): 429 - 432.
[4] Laffer CL, Scott RR, Titze JM, et al. Hemodynamics and salt-and-water balance link sodium storage and vascular dysfunction in salt-sensitive subjects [J]. Hypertension, 2016, 68(1): 195 - 203.
[5] Iwamoto T, Kita S, Zhang J, et al. Salt-sensitive hypertension is triggered by Ca2+entry via Na+/Ca2+exchanger type-1 in vascular smooth muscle [J]. Nat Med, 2004, 10(11): 1193 - 1199.
[6] Machnik A, Neuhofer W, Jantsch J, et al. Macrophages regulate salt-dependent volume and blood pressure by a vascular endothelial growth factor-C-dependent buffering mechanism [J]. Nat Med, 2009, 15(5): 545 - 552.
[7] Guessous I, Bochud M, Theler JM, et al. 1999-2009 Trends in prevalence, unawareness, treatment and control of hypertension in Geneva, Switzerland [J]. PLoS One, 2012, 7(6): e39877.
[8] Schiffrin EL. Mechanisms of remodelling of small arteries, antihypertensive therapy and the immune system in hypertension [J]. Clin Invest Med, 2015, 38(6): E394 - E402.
[9] Moore KL, Stults NL, Diaz S, et al. Identification of a specific glycoprotein ligand for P-selectin (CD62) on myeloid cells [J]. J Cell Biol, 1992, 118(2): 445 - 456.
[10] Sako D, Chang XJ, Barone KM, et al. Expression cloning of a functional glycoprotein ligand for P-selectin [J]. Cell, 1993, 75(6): 1179 - 1186.
[11] Rudemiller N, Lund H, Jacob HJ, et al. CD247 modulates blood pressure by altering T-lymphocyte infiltration in the kidney [J]. Hypertension, 2014, 63(3): 559 - 564.
[12] Muller WA. Leukocyte-endothelial cell interactions in the inflammatory response [J]. Lab Invest, 2002, 82(5): 521 - 533.
[13] 曾宪录, 王晓光, 高艳光, 等. PSGL-1——一种介导白细胞粘附的重要分子 [J]. 分子科学学报, 2002, 18(2): 84 - 89.
[14] Tinoco R, Otero DC, Takahashi AA, et al. PSGL-1: a new player in the immune checkpoint landscape [J]. Trends Immunol, 2017, 38(5): 323 - 335.
[15] Frenette PS, Denis CV, Weiss L, et al. P-Selectin glycoprotein ligand 1 (PSGL-1) is expressed on platelets and can mediate platelet-endothelial interactionsinvivo[J]. J Exp Med, 2000, 191(8): 1413 - 1422.
[16] Chen L, Song H, Wang Y, et al. Arterial α2-Na+pump expression influences blood pressure: lessons from novel, genetically engineered smooth muscle-specific α2 mice [J]. Am J Physiol Heart Circ Physiol, 2015, 309(5): H958 - H968.
[17] Mason RM. Fell-Muir lecture: Connective tissue growth factor (CCN2) — a pernicious and pleiotropic player in the development of kidney fibrosis [J]. Int J Exp Pathol, 2013, 94(1): 1 - 16.
[18] Jose PA, Yang Z, Zeng C, et al. The importance of the gastrorenal axis in the control of body sodium homeostasis [J]. Exp Physiol, 2016, 101(4): 465 - 470.
[19] Fujita T. Mechanism of salt-sensitive hypertension: focus on adrenal and sympathetic nervous systems [J]. J Am Soc Nephrol, 2014, 25(6): 1148 - 1155.
[20] Muller WA. Leukocyte-endothelial cell interactions in the inflammatory response [J]. Lab Invest, 2002, 82(5): 521 - 533.
[21] Grobe JL, Buehrer BA, Hilzendeger AM, et al. Angiotensinergic signaling in the brain mediates metabolic effects of deoxycorticosterone (DOCA)-salt in C57 mice [J]. Hypertension, 2011, 57(3): 600 - 607.
[22] Kirabo A, Fontana V, de Faria AP, et al. DC isoketal-modified proteins activate T cells and promote hypertension [J]. J Clin Invest, 2014, 124(10): 4642 - 4656.
[23] 刘力生. 中国高血压防治指南2010 [J]. 中国医学前沿杂志(电子版), 2011, 3(5): 42 - 93.
[24] Thang LV, Demel SL, Crawford R, et al. Macrophage depletion lowers blood pressure and restores sympathetic nerve α2-adrenergic receptor function in mesenteric arteries of DOCA-salt hypertensive rats [J]. Am J Physiol Heart Circ Physiol, 2015, 309(7): H1186 - H1197.
[25] Sprague AH, Khalil RA. Inflammatory cytokines in vascular dysfunction and vascular disease [J]. Biochem Pharmacol, 2009, 78(6): 539 - 552.
[26] Dmitrieva NI, Burg MB. Secretion of von Willebrand factor by endothelial cells links sodium to hypercoagulability and thrombosis [J]. Proc Natl Acad Sci U S A, 2014, 111(17): 6485 - 6490.
[27] Muller WA. Leukocyte-endothelial cell interactions in the inflammatory response [J]. Lab Invest, 2002, 82(5): 521 - 533.
[28] 赵譞, 杨新春, 蔡军. T细胞免疫与高血压的研究进展 [J]. 心血管病学进展, 2012, 33(1): 28 - 31.
[29] Krishnan SM, Dowling JK, Ling YH, et al. Inflammasome activity is essential for one kidney/deoxycorticosterone acetate/salt-induced hypertension in mice [J]. Br J Pharmacol, 2016, 173(4): 752 - 765.
[30] Mehaffey E, Majid D. Tumor necrosis factor-α, kidney function, and hypertension [J]. Am J Physiol Renal Physiol, 2017, 313(4): F1005 - F1008.
[31] Mattson DL. Infiltrating immune cells in the kidney in salt-sensitive hypertension and renal injury [J]. Am J Physiol Renal Physiol, 2014, 307(5): F499 - F508.
[32] Ramseyer VD, Garvin JL. Tumor necrosis factor-α: regulation of renal function and blood pressure [J]. Am J Physiol Renal Physiol, 2013, 304(10): F1231 - F1242.
[33] Kular L, Pakradouni J, Kitabgi P, et al. The CCN family: a new class of inflammation modulators? [J]. Biochimie, 2011, 93(3): 377 - 388.
[34] Abreu JG, Ketpura NI, Reversade B, et al. Connective-tissue growth factor (CTGF) modulates cell signalling by BMP and TGF-β [J]. Nat Cell Biol, 2002, 4(8): 599 - 604.
[35] 黄平, 钱康. 绞股蓝总皂苷对糖尿病肾病大鼠结缔组织生长因子表达的影响 [J]. 中华中医药学刊, 2007, 25(11): 2235 - 2238.
[36] Liu SC, Hsu CJ, Chen HT, et al. Correction: CTGF increases IL-6 expression in human synovial fibroblasts through integrin-dependent signaling pathway [J]. PLoS One, 2015, 10(12): e144569.
