樟树长链脂肪酰基CoA合成酶基因9克隆与表达分析
2018-05-30汪信东章挺杨海宽郑永杰江香梅
汪信东 章挺 杨海宽 郑永杰 江香梅


摘 要:該研究以樟树转录组数据为基础,筛选克隆了拟南芥AtLACS9同源候选基因CcLACS9,二者序列相似性为75%。相关软件预测CcLACS9享有植物LACS亚家族成员3个特征motifs,且N端含有定位质体信手肽。在Δlacs缺陷型酵母互补测试中,以油酸作为唯一外源脂肪酸、转化了CcLACS9的突变型酵母恢复正常生长,证明CcLACS9具有典型的脂肪酰基CoA合成酶的功能。为探究CcLACS9是否参与了樟树籽油生物合成,进一步研究了其组织表达模式和在种子发育过程中其表达量与籽油累积量之间的关系。实时荧光定量PCR分析显示CcLACS9基因在种仁与花中优势表达,种仁中相对表达量是根中的17.74倍。随机测定了30棵成年樟树成熟期种子千粒重、籽油含量和中链脂肪酸比例等指标。根据仁油含量将测试群体划为高、中、低三个不同品级,并在各品级中挑选3棵单株、逐月关联分析其仁油含量与CcLACS9相对表达量。结果表明:在种仁发育前期,仁油含量和CcLACS9表达量都持续上升且二者呈正相关性,8月份为CcLACS9表达量峰值期;9月下旬后,仁油含量趋向稳定但CcLACS9表达量仍处于较高水平但呈现下降趋势,二者无明显相关性。LACS亚家族在植物进化中较为保守,同源基因在不同植物中具有相同或相似的功能。该研究结果暗示CcLACS9可能拥有AtLACS9相似的生物学功能,即在樟树种仁油酯合成和累积过程中起重要作用。
关键词:樟树,长链脂肪酰基CoA合成酶9,籽油,基因表达分析,酵母互补检测
中图分类号:Q943
文献标识码:A
文章编号:1000-3142(2018)10-1335-11
Abstract:In this study,the cDNA encoding AtLACS9 homologous gene was identified and cloned based on transcriptomes data of Cinnamomum camphora,sharing 75% sequence similarity to AtLACS9 and being registered to CcLACS9. Multiple sequences alignment showed CcLACS9 possessed three plant LACS-specific motifs and plastidical targeting signal in N-terminal. Using oleic acid as a substrate,CcLACS9 could activate free fatty acids into acyl-CoA thioesters in a yeast mutant deficient in LACS complementation test. The tissue-specific expression profile further revealed that CcLACS9 was predominantly expressed in developing seeds and flowers,but fewer in leaf and stem by real-time quantitative PCR analysis. There were 17.74-folds relative quantitative expression of CcLACS9 in kernels relative to roots. In an effort to better understand whether CcLACS9 involved seed oil biosynthesis in camphor kernel,the correlation between expression of CcLACS9 and seed oil content was surveyed. The seeds of thirty adult camphor trees were randomly sampled in November and thousand seed weight,seed oil content and the percentage of decanoic acid and lauric acid were tested and counted. According to seed oil content,the 30 individuals were subdivided into three groups and three representative plants were selected from each group to be subjected to association analysis between seed oil yield and CcLACS9 expression level in developing kernel. The results of three groups all showed that both the contents of seed oil and expression level of CcLACS9 continued to rise from June to August,and there were significant positive correlations between them. The peak of expression level of CcLACS9 in kernel was found in August. After September,the contents of seed oil tended to be stable,however,CcLACS9 keeped high expression level and had no correlation with contents of seed oil. In plant,the homoeologous genes of LACSs subfamily tend to carry the common function. These resluts implies that CcLACS9 possibly play an important role in seed oil accumulation in kernel of camphor tree.
Key words:Cinnamomum camphora,long chain fatty aycl-coA synthetase 9,seed oil,gene expression analysis,yeast complementation
樟树(Cinnamomum camphora)是我国潜在可持续开发能源树种之一,种仁中油脂占其自身干重的55%~65%,主要以癸酸和月桂酸为主、比例在90%以上(赵曼丽等,2012)。癸酸(C10)和月桂酸(C12)属于中链脂肪酸,具有凝固点低、氧化稳定性好、可被迅速吸收、相容性和延展性佳等特性,廣泛应用于工业、医药、保健、化妆、动物养殖等行业(宫雪等,2012;刘梦芸等,2016)。樟树广布于南方诸省,据统计仅湖南一省每年产籽油便可超过400 t,作为我国鲜有的富含中链脂肪酸植物资源具有良好的开发潜力。然而,目前樟树籽油未被规模化开发利用,缺少具有竞争力的良种是其关键因素之一。作为高杂合的木本植物,樟树个体间籽油含量和成分差异显著,如何为良种定向筛选和培育提供约束性理论指标显得尤为重要。加强樟树油脂生物合成研究、特别是关键基因挖掘,填补相关基础研究空白,将有助于今后良种培育和樟树油脂产业开发。
长链脂肪酰基CoA合成酶(long chain fatty aycl-coA synthetases,LACSs)可催化游离脂肪酸(C14~C20)形成脂肪酰基CoA参与生物体内各类脂类代谢反应,在植物发育、脂肪酸延伸、植物种子油脂即三酰基甘油TAG(triacylglycerol)形成和β-氧化、生物膜合成与细胞信号转导等生物过程中具有重要作用(Watkins,1997; Khurana et al,2010)。在高等植物中,LACS是一类序列保守的蛋白,具有腺苷合成酶基因超家族(adenylate-forming enzymes superfamily,AAE)的共同特征,即包含一段高度保守的motif1(T[SG]-S[G]-G-[ST]-T[SE]-G[S]-X-P[M])和motif2(Y[LWF]-G[SMW]-X-T[A]-E)组成(注X代表任意氨基酸)的AMP-绑定结构域和享有相似的催化反应机制:第一步在消耗ATP的条件下形成酰基-AMP中间体、同时释放焦磷酸盐;第二步,将酰基转移至最终受体并释放AMP(Babbitt et al,1992; Stuible et al,2000)。AMP-绑定结构域是第一步反应的主要执行者,也是腺苷合成酶基因超家族成员鉴定的特征探针序列(Stuible et al,2000)。LACSs包含的另一段由约25个高度保守氨基酸残基组成的保守结构域称为酰基CoA合成酶(ACS)信号序列,可能是脂肪酸的结合部位和 ACS 的激活位点(Mashek et al,2007)。此外,一段由45~70个不等残基组成的专一连接区域(linker)则是LACSs区别于其它ACS家族成员的特征序列,其为LACSs成员行使正常生物学功能所必需(Steinberg et al,2000; Iijima et al,1996)。
植物LACSs亚家族各成员功能研究在模式植物拟南芥中的开展较为深入和广泛。在拟南芥中已鉴定获得9个编码LACSs基因,缺陷型酵母互补测试证实了当中7个编码蛋白都具有较强的脂酰CoA合成酶活性,而在体外催化测试中则9个编码蛋白皆具有较强的活性、但底物偏好性各异(Shockey et al,2002)。表达分析显示,除AtLACS5(花中特异表达)之外、其它成员在各组织中普遍转录但具有明显器官或组织特异性(Shockey et al,2002)。AtLACSs亚家族成员具有不同的亚细胞定位,在脂肪酸相关油脂代谢不同节点上起着重要作用,对植物正常各器官组织发育亦至关重要(Fulda et al,2002; L et al,2009; Schnurr et al,2004; Jessen et al,2011)。其中,AtLACS9基因在发育种子和莲座叶中优势表达,其编码蛋白(At1g77590)定位于质体膜上(Schnurr et al,2002)。尽管T-DNA插入未引起lacs9-1突变体发育和表型明显变化,但叶绿体中长链脂肪酰基CoA合成酶活性只维持约为野生对照个体的10%,表明AtLACS9是质体中主要的长链脂肪酰基CoA合成酶类但存在另一种LACSs与之共同介导脂肪酸运出质体(Schnurr et al,2002)。随后,双突变鉴定证实了AtLACS9与AtLACS1功能存在部分冗余,在籽油合成过程中起重要作用(Zhao et al,2010)。
1 材料与方法
1.1 材料与主要试剂
所用材料取自江西省林业科学院院内10年生至20年生的小样本樟树群体。樟树LACS9基因序列参考国家林业局樟树工程技术研究中心构建的樟树五种化学类型(芳樟醇型、桉叶油素型、樟脑型、龙脑型与异橙花椒醇型)叶组织转录组数据(江香梅等,2014)。
RNA提取试剂盒(RNAiso for Polysaccharide-rich Plant Tissue)、TaKaRa LA Taq、反转录试剂盒(First Strand cDNA Synthesis Kit)、荧光定量试剂盒(SYBR Green I qPCR Kit)、琼脂糖凝胶回收试剂盒及PMD18-T克隆试剂盒均购自 TaKaRa公司。酵母表达载体pYES2和缺陷型酵母(Saccharomyces cerevisiae)YB525菌株由江苏大学谭小力教授惠赠。各链长脂肪酸购于Sigma公司。常规试剂和培养基购自上海生工;克隆测序由上海生工完成。
1.2 方法
1.2.1 小样本樟树群体种子千粒重和种仁含油量和成分调查 随机挑选30棵长势相仿的樟树,于2015年11月采集种子2 kg,洗净、晾干,利用千分之一克电子天平称量、统计千粒重。剥取种仁、阴干后,称取100 g种仁采用索氏提取法提取油脂。称取油脂重量,计算单株籽油出油率;通过气相-质谱联用(GC-MS)测定籽油中各中链脂肪酸成分和相对含量。
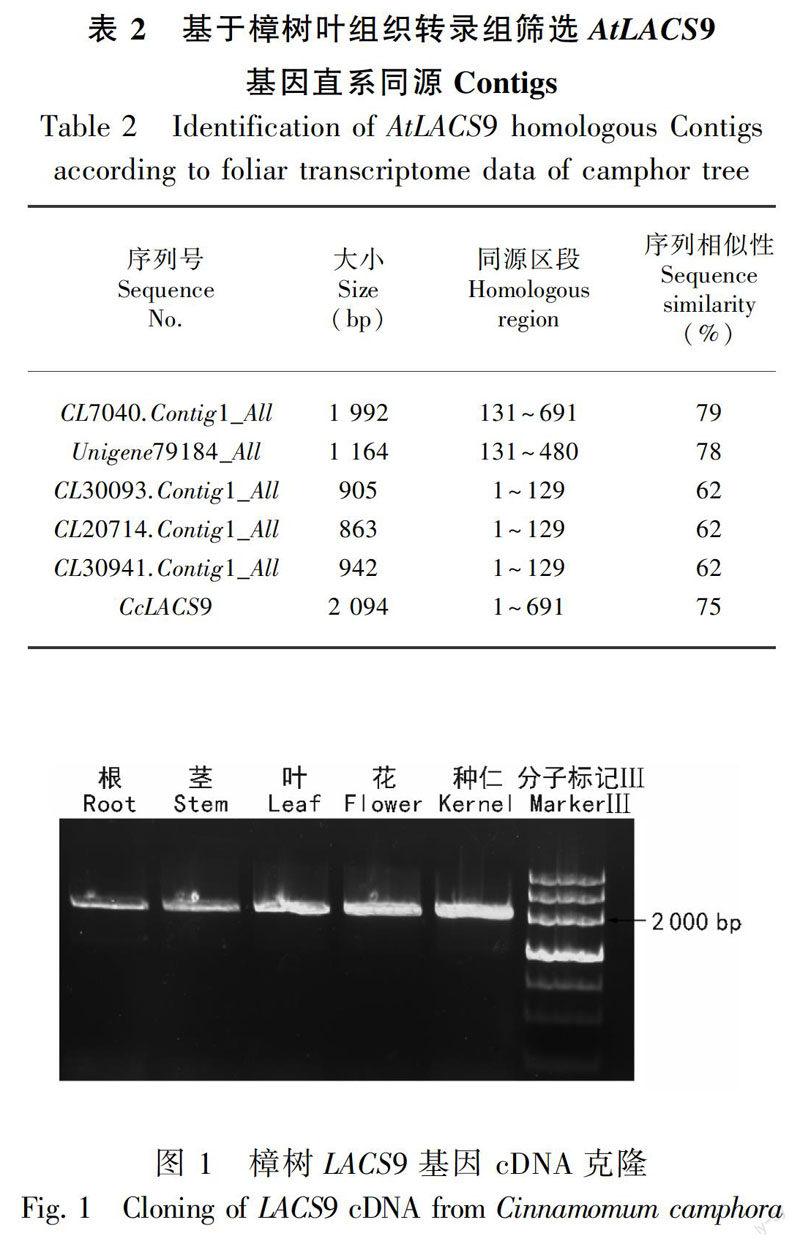
1.2.2 樟树候选LACS9基因筛选 查询Pfam(http://pfam.xfam.org/)数据库、下载AAE家族特征结构域的隐马氏模型文件(Pfam号码:PF00501)(Conti et al,1996),运行HMMER3.0程序(http://hmmer.janelia.org/)注释樟树五种化学类型叶组织转录组序列;根据注释结果,筛选获得樟树转录组中注释为AAE超家族的全部Contigs序列;以AtLACS9作为参考序列建库,运行本地blast X,比对樟树AAE超家族候选Contigs序列,根据相似性挑选AtLACS9同源Contigs;运行序列拼接软件CAP3,完成樟树LACS9基因电子克隆。
1.2.3 樟树LACS9基因克隆 于2016年4—7月,分别采集樟树茎、叶、根、花和种仁五种组织,速冻于液氮中。根据TaKaRa公司RNAiso for Polysaccharide-rich Plant Tissue说明书提取上述五种组织total RNA。DNAase I消化后去除残留DNA,取50 ng total RNA,使用PrimeScript 1st Strand cDNA Synthesis Kit反转录试剂盒合成cDNA第一链。参考由樟树五种化学类型叶组织转录组数据提供的LACS9基因电子克隆序列、设计特异引物,以樟树上述五种组织cDNA为模板,采用PCR法特异扩增樟树LACS9基因。特异引物为F:5′-TTGCGAGAAATGGCTGAAT-3′;R:5′-AAGTTCCAACCAACGGATTPCR-3′。反应体系总体积为20 μL,包括LA Taq 0.2 μL、 cDNA 1 μL、 10×LA Taq buffer 2 μL、上下游引物各0.5 μL(10 μmol·L-1)、dNTPs 1 μL(2.5 mmol·L-1)和ddH2O 14.8 μL。PCR反应程序:95 ℃预变性3 min;95 ℃ 30 s,56 ℃ 30 s,72 ℃ 2 min;72 ℃ 10 min;35个循环。1%琼脂糖凝胶电泳检测PCR产物,切取与目标产物大小相当的电泳条带,回收后与PMD18-T克隆载体连接,42 ℃热激转化大肠杆菌TOP10感受态细胞。菌液PCR筛选阳性克隆,送至上海生工测序。
1.2.4 樟树LACS9基因序列分析与鉴定 使用Scanprosite Results viewer 工具搜索注释的樟树LACS9保守域信息;使用Clustalx软件进行多序列比对,计算樟树LACS9基因与其它植物直系同源基因序列相似性;使用SignalP 4. 1 Server和Predictprotein等软件预测樟树LACS9亚细胞定位;引入擬南芥、油菜、花生等植物LACS9,通过MEGA6.0软件分析植物LACS9基因谱系发生关系。
1.2.5 樟树LACS9基因表达组织特异性分析 于2016年4—7月,分别采集樟树花、茎、叶、种仁和根组织,提取total RNA。以Actin基因作为内参对照基因(F:5′-CCTCGACACACAGGCGTTAT-3′;R:5′-CCATGCTCGATGGGATATTTCA-3′),采用实时荧光定量PCR方法(qRT-PCR)检测樟树LACS9基因在樟树不同组织中表达情况。qRT-PCR引物:F:5′-ACCTGCCTTTGGCTCACA-3′,R:5′- AAGGCGATCCGTATCCAA-3′。PCR反应在Bio-RAD C1000 TM Thermal Cycler 荧光定量PCR仪上完成。 反应体系总体积20 μL,包括(cDNA 50 ng,2 × SYBR Green qPCR10 μL,10 mmol·L-1的正反向引物各1 μL,超纯水补足至20 μL),反应程序:95 ℃ 2 min; 95 ℃ 15 s,58 ℃ 30 s,72 ℃ 20 s;40个循环,添加溶解曲线;每个样品重复3次。实验数据统计参照2-△△CT计算相对表达量(Livak & Schmittgen,2001)。
1.2.6 樟树种仁发育过程中LACS9基因表达量与发育进程中种仁油脂含量关联分析 根据上年种仁油测定结果,将测试小样本分为三个品级,即出油率>60%为高品级、出油率介于50%~60%为中品级和出油率<50%为低品级。每个品级随意选取3棵作为代表植株,于2016年5—11月,逐月采集樟树种子、剥取种仁。一部分种仁用于提取籽油,测定油脂含量和成分;一部分种仁用于总RNA提取,测定樟树种子发育各时期
LACS9基因相对表达量。籽油提取采用索氏提取法并通过GC-MS测定含量和成分。种仁总RNA提取和LACS9基因实时荧光定量表达分析同上。采用SPSS Statistics 19.0统计软件分析种子发育过程中LACS9基因表达量和油脂含量之间的关联性。
1.2.7 樟树LACS9缺陷型酵母互补实验 酵母菌株YB525(faa1Δfaa4Δ)缺少活化外源脂肪酸所必需的LACSs,不能在基本培养基中正常生长,常用于外源LACSs活性分析(朱福各等,2009)。将重组质粒pYES2-CcLACS9和对照质粒pYES2分别转化至缺陷型酵母YB525感受态细胞中,通过缺省尿嘧啶固体培养基筛选阳性转化克隆。随机挑取阳性克隆,在缺省尿嘧啶液体培养基中培养至对数期中后期,低速离心收集酵母细胞,加入2 mol·L-1的山梨醇、漂洗两次。将细胞转接至缺省尿嘧啶液体培养基(含2%半乳糖且不含葡萄糖)中,振荡培养4~5 h、以诱导外源目标基因大量表达。按1%比例吸取菌液加入到缺省尿嘧啶液体培养基(含2%半乳糖和0.1% Tritonx-100)中,并添加98 μmol·L-1的脂肪酸C18∶1作为外源脂肪酸。于30 ℃振荡培养约84 h,各取1 mL培养基利用分光光度计测量菌体密度来衡量其生长速度。
2 结果与分析
2.1 小样本樟树群体种子千粒重和种仁含油量调查
2015年11月采集的30棵樟树种子千粒重最高值为184.23 g,最低值为118.62 g,平均值为149.92 g。在测试小样本中,2/3樟树个体种子千粒重在140~180 g之间,方差为16.92,呈类“正态”分布。测试样本出油率最高为76.78%,最低为42.48%,均值为57.37%。出油率高于60%的有15株,低于50%的有12株,介于50%~60%之间的有3株,分布呈“两极化”。脂肪酸成分分析表明,测试樟树种仁中中链脂肪酸相对含量平均为95.71%,除个别单株外都介于94%~97%之间,分布较集中。中链脂肪酸当中癸酸含量(平均为62.36%)显著高于月桂酸含量(平均为33.35),二者呈负相关性(r=-0.86,P<0.01)。在5个调查指标中,方差值表现为千粒重>出油率>癸酸相对含量>月桂酸相对含量>大于中链脂肪酸相对含量。
2.2 樟树LACS9基因筛选与cDNA序列克隆
在樟树叶组织转录组数据中搜索到5条Unigenes或Contings序列(表1),可能为拟南芥LACS9基因直系同源序列。借助软件CAP3对5条Unigenes或Contings序列进行拼接,获得了包含完整开放阅读框的樟树LACS9基因电子克隆。根据电子克隆设计引物,分别以樟树茎、叶、根、花和种仁五种组织cDNA为模板、通过PCR法进行全长cDNA序列特异扩增,1% 琼脂糖凝胶电泳检测获得了清晰的单一条带(图1)。经单克隆测序显示,扩增条带大小为2 356 bp,包含2 094 bp的编码区序列、编码697个氨基酸多肽,命名为CcLACS9。EditSeq软件预测显示,CcLACS9大小为86.83 kDa,等电点为7.39、生理中性条件下略偏碱性。ProtParam软件在线分析显示CcLACS9含有多个较强的亲水区域,为亲水性蛋白。提交GeneBank数据库,获得登入号MF966481。
2.3 樟树LACS9序列鉴定及进化分析
AMP绑定结构域(PROSITE PS00455)[LIVMFY]-{E}-{VES}-[STG]-[STAG]-G-[ST]-[STEI]-[SG]-x-[PASLIVM]-[KR]是植物ACS家族的标志性标签。为了鉴定CcLACS9是否属于植物ACS家族成员,利用Scanprosite Results viewer软件分析了其保守结构域。结果显示(图2:A),在CcLACS9蛋白序列第256~267和第452~462位分别存在12个和10个氨基酸残基(图中*标记)组成的高度保守的AMP绑定结构域;第531~560位包含由30个氨基酸(图中☆标记)组成高度保守的ACS信号序列,表明CcLACS9屬于典型的植物ACS基因家族成员。此外,介于AMP绑定结构域和ACS信号序列之间,还存在一段包含71个氨基酸残基组成的保守结构域(第354~424,图中◆标记)即为LACSs亚家族特殊的linker结合域,该结合域在不同物种与不同成员中序列长度有所差异。综上所述,CcLACS9既具备 ACS 家族的共同特征,又具有真核生物 LACSs专有的linker结合域,可基本确认其为樟树所含有的LACSs亚家族成员之一。将CcLACS9与其它植物LACSs亚家族同源基因进行了多序列比对,结果显示AMP绑定结构域、ACS信号序列和linker结合域都具有极强的保守型(图2:B)。
CcLACS9与油棕(Elaeis guineensis)、莲花(Nelumbo nucifera)、无油樟(Amborella trichopoda)、二倍体棉花(Gossypium raimondii)、芝麻(Sesamum indicum)、拟南芥(Arabidopsis thaliana)、麻枫树(Jatropha curcas)、油菜(Brassica napus)、玉米(Zea mays)、蓖麻(Ricinus communis)等物种中同源基因序列相似性分别为80%、81%、80%、78%、78%、75%、78%、75%、75%和76%,表明在植物进化中LACS9极为保守,可能在不同植物中都具有相同或相似的功能。LACS9谱系发生分析显示,CcLACS9进化上介于单子叶植物和双子叶植物LACS9之间、与莲LACS9较为相似,该结果与樟树在陆生植物中的进化地位相符。SignalP 4. 1Server和Predictprotein等亚细胞定位预测软件都预测CcLACS9定位于质体,暗示其极可能与AtLACS9一样,执行了将生物合成后脂肪酸运出质体的任务且进而对种仁中油脂合成调控具有重要作用。
2.4 樟树LACS9基因表达具有组织特异性
提取樟树花、叶、茎、根和种仁等组织total RNA,以樟树Actin基因为内参、通过荧光定量PCR法分析了CcLACS9基因在不同组织中的表达情况。从图3可以看出,CcLACS9基因在花、叶、茎、根和种仁中均有表达,但表达丰度具有明显的差异性。其中,CcLACS9基因在花、种仁中优势表达最为明显,叶组织次之,在三者中表达量远高于其它组织。将根中CcLACS9基因表达量设定为1,在茎、叶、花和发育种仁等四个组织中相对表达量分别是其 4.75、9.31、15.82和17.74倍。
2.5 樟树种仁发育前期油脂含量与CcLACS9基因表达量正相关
根据种仁含油量,将供试樟树小样本群体划分为上(>60%)、 中(50%~60%)和下(<50%)三个品级。每个品级随机抽样3棵樟树进行各发育时期种仁含油量分析。2016年6—11月逐月采集樟树种子、剥取种仁,阴干后用于油脂提取。从图6可以看出,樟树种子发育前期(6—8月),随着种子个体不断发育膨大、种仁油脂含量(出油率)呈快速增长态势;9—11月,随着种子逐渐发育至成熟阶段,种仁中油脂含量增长缓慢至趋于停滞。各品级樟树油脂含量在8月开始呈现显著差异并保留至成熟期(11月),所有单株不同脂肪酸比例在整个发育时期大体稳定。
对应地,我们通过荧光定量PCR方法测定了各品级樟树种仁发育过程中CcLACS9基因的变化情况,以及研究该基因表达量与种仁含油量之间的关系。从图6可以看出,相比较于5月初种子刚出现时期,6—11月时期种仁中CcLACS9基因都呈现出显著的上调表达态势,其中表达高峰出现在8月份。同一时期,CcLACS9基因相对表达量在不同品级各单株间无明显的差异且具有相似的时空表达模式。应用SPSS Statistics 19.0软件分析二者关联情况,结果表明在种子发育前期(6—8月)、种仁含油量与CcLACS9基因相对表达量呈现明显正相关性(r=0.96);后半期(9—11月)、种仁含油量与CcLACS9基因无明显的相关性(r=0.13),但二者都维持高水平。
2.6 樟树LACS9可恢复缺陷型酵母YB525正常生长
通过酶切连接的方法将CcLACS9基因CDS区插入表达载体pYES2多克隆位点,重组质粒通过酶切鉴定和测序验证正确。将对照原始质粒pYES2和重组质粒pYES2-CcLACS9分别转化缺陷型酵母YB525菌株,挑选阳性转化子培养于含有C18∶1(油酸)作为唯一碳源的缺省尿嘧啶液体培养基中,28 ℃振荡培养至对数期中期(约84 h)。OD600值测定显示,转化了pYES2-CcLACS9重组质粒的缺陷型酵母细胞可以正常生长,而转化了原始pYES2质粒的转化子不能生长。实验结果表明 CcLACS9能够互补酵母缺陷型,证明其具有脂酰CoA合成酶活性(图7)。
3 讨论
据调查,樟、沉水樟、毛豹皮樟、山橿、大叶木姜子、舟山木姜子、海南木姜子等多种樟科植物种仁平均含油量超过50%,是我国潜在的重要的生物质能源开发树种之一(祝必琴等,2014)。然而,目前相关资源还未获得有效利用与开发,樟树大量种子脱落甚至作为行道绿化时的“负产物”而难以处理,缺乏有竞争力的良种是其主要原因之一,需要开展针对性的高含油量的品种选育。在本研究所测试的30棵小样本樟树群体中,单株间种子千粒重和种仁出油率差异明显,差异极值分别为65.61克和34.3%,表明在自然界中樟树油脂良种筛选中具有很大的选育空间。千粒重和出油率之间无明显相关性,在选育过程中二者应综合被考虑。
植物脂肪酸从头合成始于质体中,在脂肪酸合成酶催化下以丙二酰-ACP和乙酰-CoA作为起始底物进行连续聚合反应,以每个循环增加2个碳的频率延伸酰基碳链直至合成16~18个碳的饱和脂肪酰-ACP(Brown et al,2006)。依次在Δ9硬脂酰去饱和酶和酰基-ACP水解酶作用下形成游离非饱和脂肪酸(Kachroo et al,2007)。为完成诸如三酰甘油、蜡质、角质及软木质等物质合成,一大部分游离非饱和脂肪酸需以酰基-CoA激活形式运出质体,该反应过程由LACSs介导完成。实验数据表明AtLACS9在拟南芥种仁油脂合成和累积过程中起重要作用(Schnurr et al,2002)。在本研究中,CcLACS9基因序列与AtLACS9基因相似性为75%,亚细胞定位预测其定位于质体中且在种子中优势表达,暗示其极可能作为AtLACS9直系同源基因在樟树籽油合成与累积过程中起重要作用。4月底樟树开始挂果,5—8月是果实快速膨胀期和籽油合成累积高峰期,9月之后逐渐过渡到成熟期。CcLACS9在种子发育过程前期(5—8月)处于持续上调表达状态,其表达量与种仁油脂含量呈正相关,再次暗示了其在樟树籽油合成和累积过程中起了重要作用。种子成熟后(9月后),CcLACS9表达量与种仁油脂含量无明显相关性,其维持较高表达水平可能主要在于弥补呼吸作用等引起的能量消耗、维持种仁油脂含量动态平衡。不同单株中,CcLACS9基因表达量差异性与油脂含量差异性无明显相关性,暗示还有其它因素参与了籽油合成调控。
樟树籽油富含中链脂肪酸成分,是研究中链脂肪酸生物合成机制的好材料。酰基-ACP水解酶是调控樟树合成中链脂肪酰基的关键基因之一(Yuan et al,1995),然CcLACS9激活游离脂肪酸是否具有链长选择性还未可知。在本研究中,我们特意比较了UFA(unusual fatty acid)植物包括樟树、油棕和蓖麻与普通植物如拟南芥、大豆、芝麻等LACS9序列特征,结果显示植物LACS9为一类较保守的LACSs亚家族成员,所有物種间该同源基因序列相似性介于75%~81%之间,且无明显差异性motifs存在。利用MEGA6.0构建植物LACS9谱系发生树,结果显示CcLACS9系统发育介于油棕、玉米等单子叶植物LACS9和芝麻、油茶等双子叶植物LACS9之间,恰与樟树作为基底被子植物的进化地位相符,来源于UFA植物的LACS9并未特异聚集在一个进化分支上。在微生物、动物相关研究中(Lindner et al,2006; Kasuya et al,2009; Meng et al,2010),中链脂肪酸酰基CoA合成酶(MACS)可专一性地“活化”中链脂肪酸。杨树MACS在体外实验中亦对己酸、壬酸和癸酸具有明显的活性(曹山等,2016)。然而,目前尚未发现定位于质体中的MACS,对其具有将植物体内中链脂肪酸转入/出质体功能存疑。樟树中是否存在特异MACS介导中链脂肪酸转入/出质体需要进一步研究。
