黄毛草莓和五叶草莓的耐热性及其对高温的响应
2018-05-30郭玲玲李钧敏
郭玲玲, 李钧敏, 闫 明
(1.山西师范大学生命科学学院,山西临汾 041004; 2.浙江省植物进化生态学与保护重点实验室/台州学院生态研究所,浙江台州 318000)
温度是植物生长和产量的一个重要限制因子[1]。随着全球气候变暖趋势愈来愈明显,高温对农业生产的影响日益引起人们的关注,由高温造成的农作物减产问题已经在番茄、葡萄、草莓等作物上得到证实[2]。草莓作为一种经济价值较高的作物,其栽培过程忌高温。有研究表明,高温条件下草莓苗的膜透性增加,溶质外渗,水分平衡失调,易造成植株萎蔫、叶绿素合成受阻、营养成分和产量下降,甚至植株死亡[3-4]。野生草莓在抗热性上优于草莓栽培种[5],其抗性因品种不同而有所差异。黄毛草莓(FragarianilgerrensisSchltdl)和五叶草莓(FragariapentaphyllaLosinsk)是2种较为常见的野生草莓,有学者认为黄毛草莓较五叶草莓有更强的耐热性[6],但对耐热性的差异原因鲜有报道。本试验以二倍体野生黄毛草莓与五叶草莓为材料,研究黄毛草莓与五叶草莓的耐热性,初步探究其耐热性差异的原因,为黄毛草莓及五叶草莓的开发利用提供理论依据。
1 材料与方法
1.1 试验材料
黄毛草莓于2012年7月采自西藏自治区林芝县;五叶草莓于同年采自四川省茂县,植株带回实验室于温室中栽培,待其活棵成壮苗,分别选择3株植株用于高温半致死温度测定。同时,取该3株植株上的匍匐茎苗定植于塑料盆中,人工气候室栽培,培养条件为光照度400 μmol/(m2·s)、相对湿度75%、光周期14 h/10 h(白天/夜晚)、温度20 ℃/15 ℃(白天/夜晚)。培养基质为泥炭土、蛭石、松树皮、沙子以体积比 2 ∶1 ∶1 ∶2 的比例混合而成,植株栽培条件基本一致,每株施以 N ∶P ∶K=13% ∶13% ∶13%的好康多缓释肥8 g/L,由匍匐茎形成的壮苗用于高温胁迫试验。
1.2 高温半致死温度的测定
参照张常隆等的方法[7]加以改进,选取黄毛草莓、五叶草莓位置相近的成熟叶片,每种草莓每株用打孔器打取9个圆片,每株各取1个圆片计3个圆片装入1个带刻度的具塞试管中,共9支试管;各加入6 mL去离子水,将其中8支试管分别置入35、40、45、50、55、60、65、70 ℃水浴中处理1 h,第9支试管直接置于室温下1 h为对照;将9支试管抽真空 15 min,25 ℃保温1 h;充分振荡,25 ℃条件下测电导率R;将试管沸水浴15 min,取出,冷却到25 ℃,充分振荡,第2次测电导率K;以室温处理的叶片测得的电导率C作为本底,计算相应温度下的相对电导率,计算公式为
Y=(R-C)/(K-C)×100%。
用Logistic方程y=k/(1+ae-bx)拟合8个处理温度与相对电导率的回归模型,参照莫慧栋的方法[8],计算出k、a、b及拟合参数r值,求出曲线出现拐点时的温度,即高温半致死温度LT50=(lna)/b。
1.3 高温胁迫试验
1.3.1 试材处理 试验于2015年8月在浙江省台州学院试验基地进行。40 ℃高温胁迫试验参照Lafta等的方法[9]经改良进行,将由匍匐茎苗形成、长势一致的植株移至人工气候室,设置人工气候室初始温度为20 ℃/15 ℃(白天/夜晚)条件下开始升温,升温速度为白天4 ℃/d、夜晚3 ℃/d[10],达到预期温度40 ℃/32 ℃(白天/夜晚)后保持温度恒定,同时,将移至人工气候室温度条件为20 ℃/15 ℃(白天/夜晚)正常栽植的草莓植株为对照;培养7 d将植株移出,测定各项指标。每处理6株,重复3次。人工气候室条件一致,光照度为 400 μmol/(m2·s)、相对湿度为75%、光周期为14 h/10 h(白天/夜晚)。
1.3.2 测定项目和方法
1.3.2.1 光合指标 选取成熟叶片的中间小叶,用美国 LI-COR 产Li-6400X型便携式光合测定系统测定净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr),测定条件设定为叶片温度25 ℃、CO2浓度400 μmol/mol、饱和光照度1 000 μmol/(m2·s);利用Regent Instrument Inc产的Win Folia叶面积分析仪扫描叶片以获得有效光合作用面积。每植株测定3次,取平均值。
1.3.2.2 相对叶绿素含量 选取成熟叶片的中间小叶,用美国产CCM-200型手持式叶绿素仪测定相对叶绿素含量。
1.3.2.3 超氧化物歧化酶(superoxide dismutase,SOD)活性及丙二醛(malomialdehyde,MDA)含量 选取成熟的新鲜叶片,分别用氮蓝四唑光还原法、硫代巴比妥酸比色法测定SOD活性、MDA含量[11-12]。
1.3.2.4 叶片远轴面表皮角质层厚度 选取成熟的新鲜叶片,在叶片中部以垂直于主脉方向将其横切制成临时装片,置于40×的中国产麦克奥迪BA410E型显微镜下观察,选靠近主脉附近的表皮细胞角质层用Motic advantages软件显微摄像,利用Image J软件测定其厚度。
1.3.2.5 叶片蜡质含量 选取成熟的新鲜叶片,用打孔器打孔获取固定面积的圆片,用加热法[13]提取蜡质,具体操作方法为称取质量为m1的叶片,将其置于预热至60 ℃的1.5 mL三氯甲烷提取液中30 s,立即取出,待三氯甲烷自然挥发,称取叶片的质量为m2,则单位面积的蜡质含量=(m1-m2)/叶片表面积。
1.3.2.6 叶片气孔相关指标 选取成熟的新鲜叶片,用打孔器打孔获取固定面积的圆片,置于70%乙醇中保存;圆片复水,置于5 mL由10%铬酸、10%硝酸铵按1 ∶1比例混合的气孔解离液中黑暗处理48 h;用镊子剥离叶片远轴面的表皮置于水中,于1%番红染液中染色2 h;倒出染液,制成临时装片,置10×BA410E显微镜下观察,用Motic advantages软件摄像,统计气孔数量,同时采用Image J软件进行显微测量获得气孔长度与宽度,每张叶片取5个视野,取其平均值,计算气孔密度、气孔开度,计算公式为:
气孔密度=5个视野下气孔个数的平均值/视野实际面积;
气孔开度=气孔宽度/气孔长度。
1.3.2.7 生物量 将草莓的地上部分、地下部分分别装入牛皮纸袋中,105 ℃杀青30 min,烘箱中75 ℃烘3 d至恒质量,分别称取地上、地下部分的生物量,计算获得根冠比。对用于其他指标测定的叶片,通过称量其鲜样质量,再换算成干质量后计入地上部分生物量。
1.4 数据处理
采用SPSS 17.0软件对数据进行分析,采用Origin 8.5软件进行作图,采用独立样本t检验分析高温处理对生理指标、叶片显微结构等的差异显著性。
2 结果与分析
2.1 黄毛草莓和五叶草莓的高温半致死温度
由图1可知,随着温度的升高,黄毛草莓、五叶草莓的相对电导率呈上升趋势。由表1可见,黄毛草莓、五叶草莓2种草莓的Logistic方程拟合系数呈极显著,高温半致死温度分别为51.43、50.70 ℃。
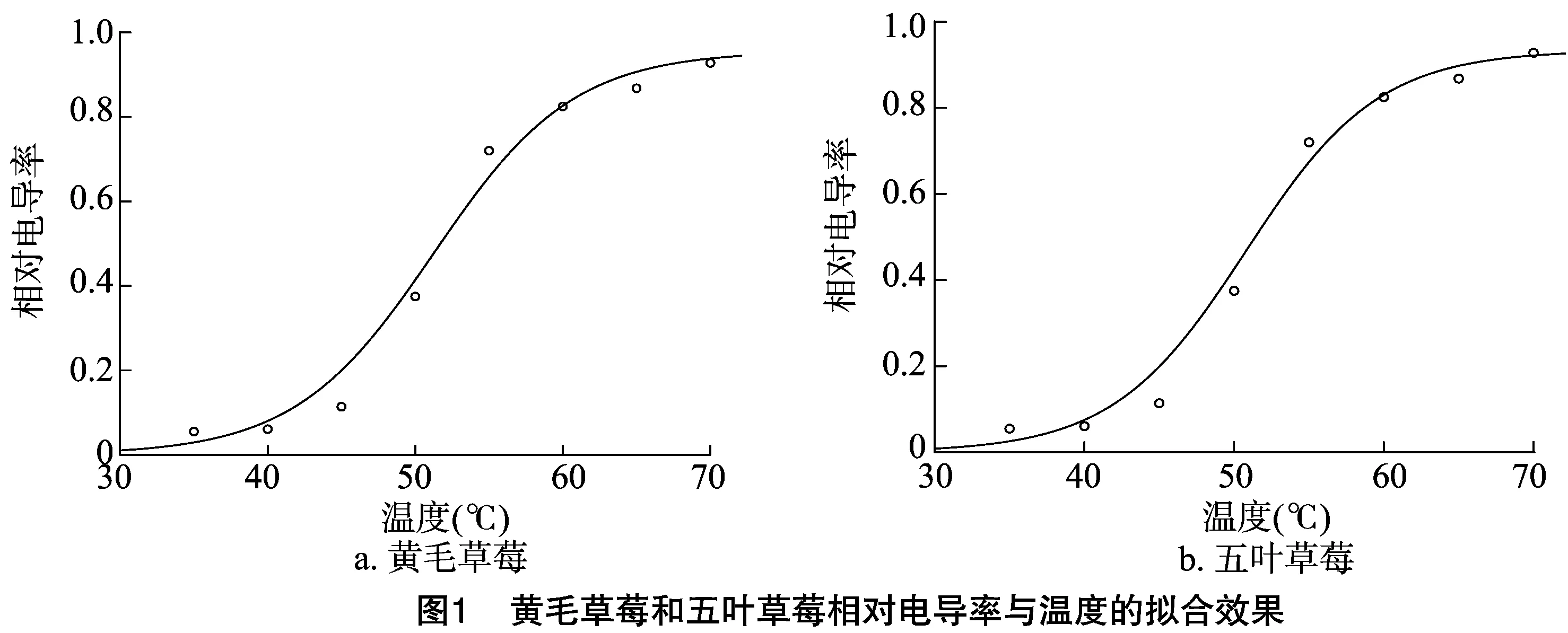

表1 黄毛草莓和五叶草莓相对电导率的Logistic方程参数及半致死温度
注:拟合系数后**表示相对电导率与温度回归相关性极显著。
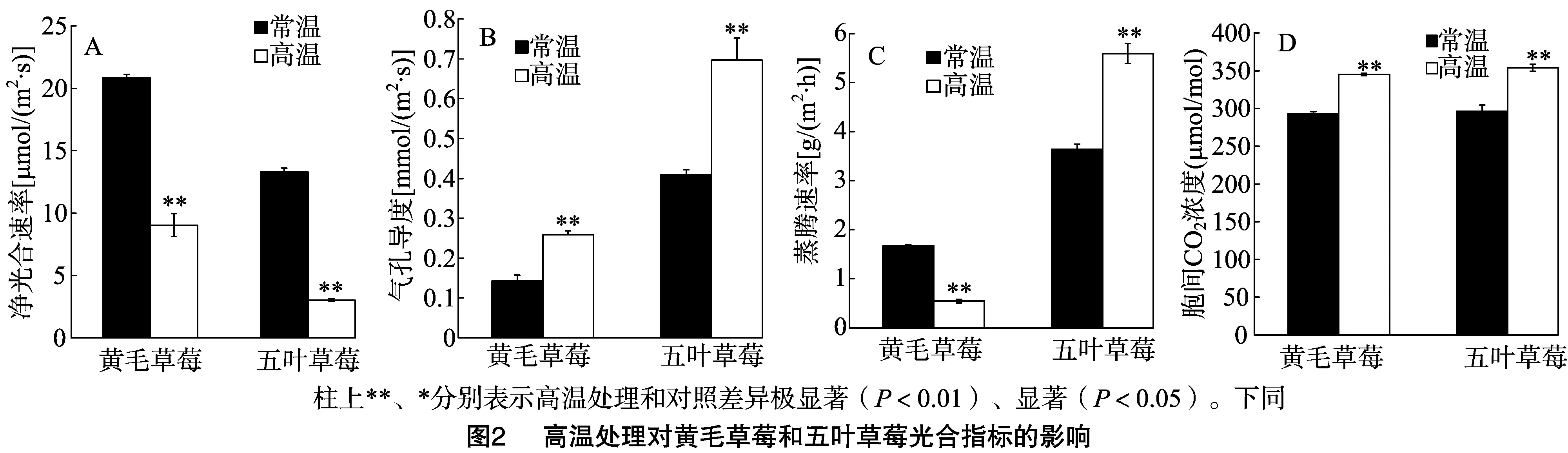
2.2 高温处理对黄毛草莓和五叶草莓光合生理的影响
由图2可知,与室温20 ℃/15 ℃(白天/夜晚)常规栽培相比,高温处理的黄毛草莓和五叶草莓其净光合速率分别极显著下降56.7%、77.2%,气孔导度分别极显著上升80.7%、41.2%,胞间CO2浓度分别极显著上升17.6%、19.5%(P<0.01),黄毛草莓蒸腾速率显著下降67.2%、五叶草莓蒸腾速率显著上升53.4%(P<0.05)。
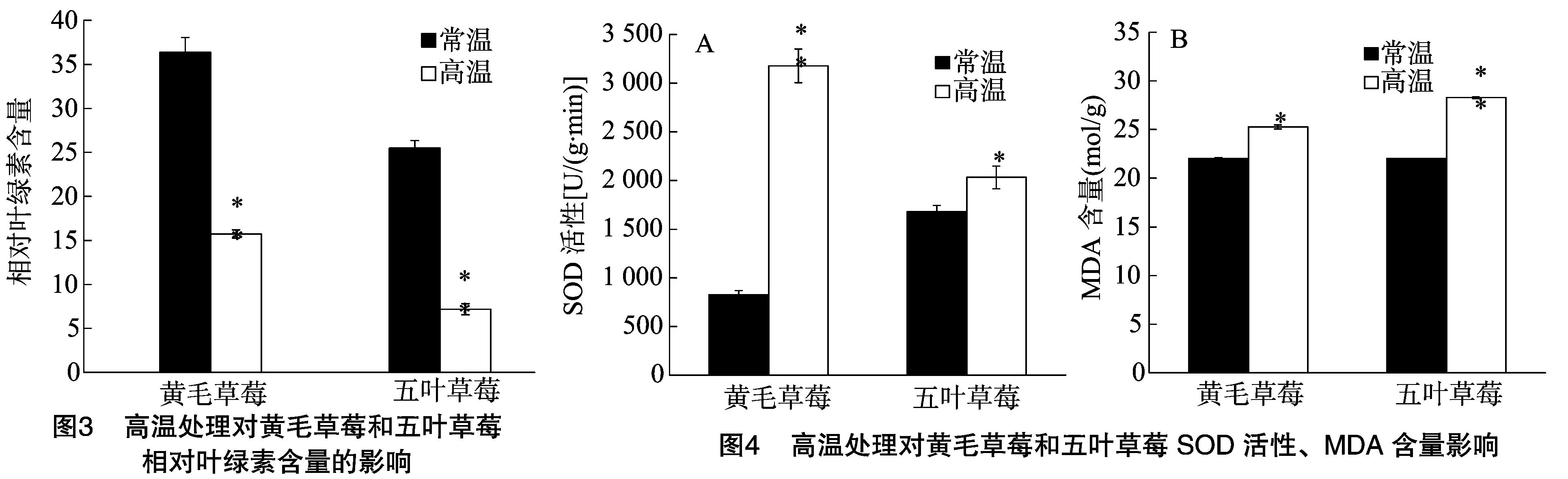
2.3 高温处理对黄毛草莓和五叶草莓相对叶绿素含量的影响
由图3可知,与对照相比,高温处理的黄毛草莓和五叶草莓相对叶绿素含量下降,分别较对照极显著降低56.7%、71.9%(P<0.01),五叶草莓相对叶绿素含量降幅大于黄毛草莓,高温处理与室温处理的黄毛草莓相对叶绿素含量相应高于五叶草莓。
2.4 高温处理对黄毛草莓和五叶草莓SOD活性、MDA含量的影响
由图4可知,与对照相比,高温处理的黄毛草莓SOD活性极显著上升284.8%(P<0.01),五叶草莓显著上升 21.9%(P<0.05),黄毛草莓SOD活性的上升幅度大于五叶草莓;高温处理的黄毛草莓SOD活性高于五叶草莓;高温处理的黄毛草莓和五叶草莓MDA含量分别上升14.8%、28.5%,五叶草莓的MDA含量增幅较大;高温处理的五叶草莓MDA含量高于黄毛草莓。
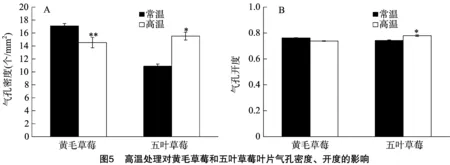
2.5 高温处理对草莓叶片气孔密度及气孔开度的影响
由图5可知,与对照相比,高温处理的黄毛草莓气孔密度有极显著下降(P<0.01),气孔开度变化不显著;五叶草莓的气孔密度和气孔开度均有显著上升(P<0.05)。
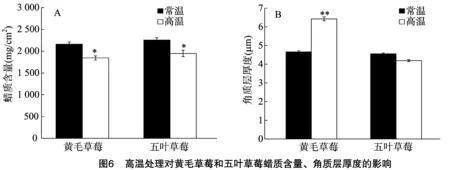
2.6 高温处理对黄毛草莓和五叶草莓叶片蜡质含量及角质层厚度的影响
由图6可知,与对照相比,高温处理的黄毛草莓和五叶草莓蜡质含量显著下降14.6%、13.6%(P<0.05),黄毛草莓的角质层厚度极显著增加37.7%(P<0.01),五叶草莓的角质层厚度略有下降,与对照差异不显著。
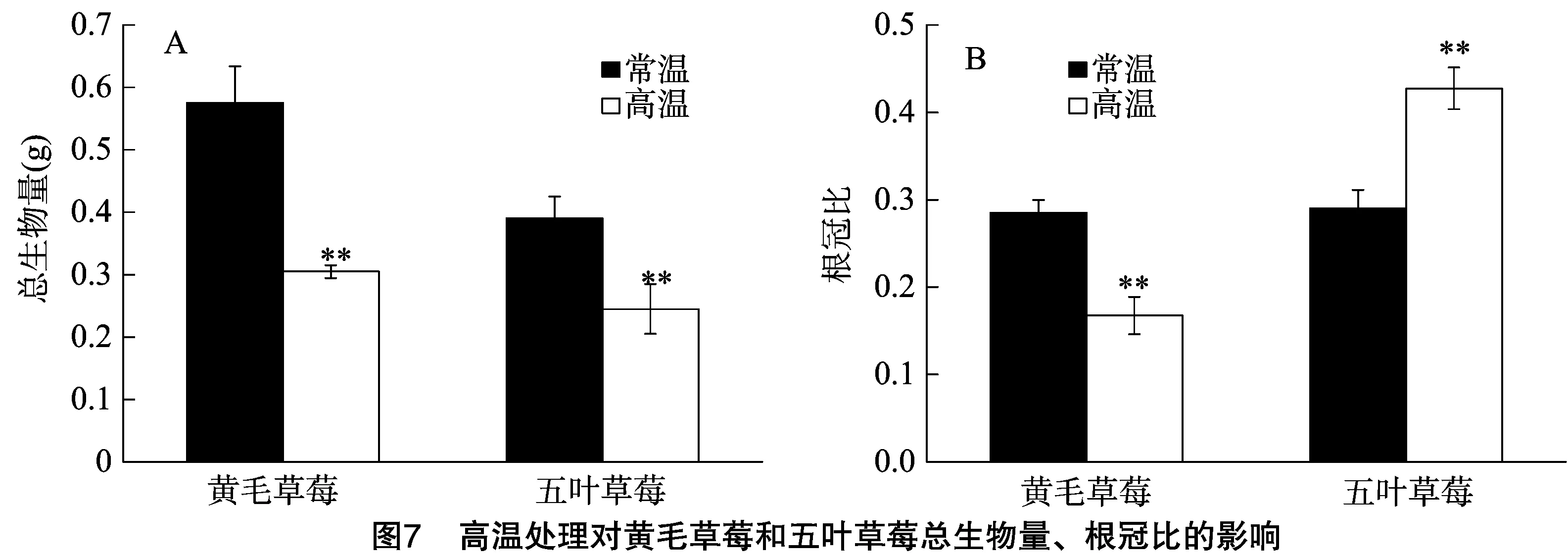
2.7 高温处理对黄毛草莓和五叶草莓总生物量、根冠比的影响
由图7可知,与对照相比,高温处理的黄毛草莓和五叶草莓总生物量有极显著降低,分别下降37.1%、47.0%,高温处理的黄毛草莓根冠比较对照极显著下降41.2%、五叶草莓极显著上升47.4%(P<0.01)。
3 结论与讨论
测定不同高温下植物叶片的电导率并拟合Logistic方程来评价植物的耐热性是最常用的方法,通过拟合曲线获得的半致死温度高低可表示植物耐热性的强弱,而半致死温度与耐高温能力往往呈正相关,夏莹莹等根据半致死温度的高低确定了4个油茶品种的耐热性顺序[14]。本研究结果显示,黄毛草莓的高温半致死温度为51.43 ℃,而五叶草莓为 50.70 ℃,表明黄毛草莓的耐热性要高于五叶草莓,这与李洪雯等的观点[6]一致。
高温对植物生长的影响很大程度上归结于光合作用的下调。试验结果表明,高温导致黄毛草莓和五叶草莓的净光合速率分别较室温处理极显著下降56.7%、77.2%,这表明前者在高温处理下净光合速率的下降幅度要小于后者,这可能是黄毛草莓具有较高耐受性的一个原因。气孔是植物体内气体交换的门户及水气蒸腾的通道,对光合作用产生明显的影响,叶片气孔开度下降会引起叶绿体内CO2供应受阻[15-16],从而导致叶片光合作用下降。气孔大小和密度变化是快速适应短期恶劣环境的策略之一[17],气孔密度与蒸腾速率、气孔扩散导度呈正相关[18]。本试验结果表明,高温处理前后黄毛草莓的气孔开度没有显著变化,而五叶草莓有显著上升;高温处理的黄毛草莓和五叶草莓气孔导度、胞间CO2浓度与净光合速率变化相反,表明高温处理下叶片光合作用的下降是由非气孔因素引起的[16];高温处理的黄毛草莓气孔密度急剧下降、五叶草莓有显著上升。黄毛草莓通过降低气孔密度来降低其蒸腾速率,以提高其对高温处理的耐受性,这与水稻突变体对高温的耐受策略[19]相似;五叶草莓通过增加气孔密度、提高气孔开度来增加其蒸腾速率,40 ℃高温处理可能会对五叶草莓带来较大损害,其通过增大气孔来增加蒸腾作用、降低植物温度,这也从侧面反映黄毛草莓对40 ℃高温的耐受性要高于五叶草莓。
植物处于高温胁迫时,叶绿体的形成和含量会受到扰乱[20]。Feierabend等证实,在大麦、玉米等5个作物中,除玉米外其他4个作物在温度超过28 ℃时叶绿素含量开始降低[20]。Mohanty等进一步分析原因认为,在胁迫条件下,叶绿素的快速降解是其含量降低的因素之一[21];杜国栋等也认为,50 ℃高温胁迫引起仁用杏叶绿素含量降低的原因是叶绿素的快速降解[16]。有研究进一步指出,耐热性强的小麦在高温下可保持较高的叶绿素含量[22]。五叶草莓经高温处理,其叶绿素含量下降幅度较黄毛草莓大,表明其叶绿素降解速率高于后者,这可能也是其耐热性相对较差的原因之一。
改变生物量分配格局是植物适应逆境胁迫的一个重要策略,生物量分配的可塑性决定植物对异质性资源的获取能力,具有重要的生态学意义[23]。干旱胁迫下,早熟禾可将更多的生长分配到根系以促进根系的生长,增加根冠比[24],而高温对植物生长的抑制会体现在生物量的降低上[25]。本试验结果表明,五叶草莓在高温处理前后生物量下降幅度大于黄毛草莓,表明其对高温处理的耐受性低于后者;40 ℃高温处理的黄毛草莓将更多的生物量分配到地上部分以促进茎、叶的生长,而高温处理带来的土壤水分胁迫由于并不是很强,可通过促进地上部分的生长来弥补光合作用下降带来的生物量损失;而五叶草莓将更多的生物量分配到地下部分以促进根的生长,高温处理已显著影响其土壤水分状况,需要更多的根系以增强其从土壤中获取更多水分的能力[26],这从侧面进一步说明黄毛草莓比五叶草莓具有更强的高温耐受性。
高温处理常会造成活性氧的积累,引起膜脂过氧化,导致丙二醛累积,从而破坏细胞膜。膜完整性的维持与植物体内的保护性酶有关,而超氧化物歧化酶(SOD)是植物的一种重要保护酶,是清除体内活性氧的第一道防线。Wahid等认为,膜完整性是植物耐热性的主要机制之一[27]。孙震认为,丙二醛含量与抗热性呈负相关[15]。本试验结果表明,高温处理的五叶草莓丙二醛含量增幅相对较大,表明其膜受到的破坏程度比黄毛草莓严重;黄毛草莓SOD活性增幅大于五叶草莓,表明黄毛草莓通过提高SOD活性来缓解膜脂过氧化的能力要高于五叶草莓,这也能一定程度上反映黄毛草莓的耐热性为何高于五叶草莓。
角质层包括蜡质和角质,是覆盖于陆生植物表面、与外界环境接触的重要保护性障碍物,在植物抵抗防止水分丢失、抵抗病菌入侵、抵抗紫外辐射和霜冻等各种生物、非生物胁迫中起着非常重要的作用[28]。40 ℃高温处理的黄毛草莓角质层厚度有显著增加,而五叶草莓无明显变化,表明高温处理的黄毛草莓通过增加角质层厚度来增强其抗逆性,这与王家训等的研究结果[29]一致。有关高温处理影响植物蜡质的相关研究较少。本研究发现,40 ℃高温处理的黄毛草莓、五叶草莓蜡质含量有显著下降,两者之间的降幅差异不显著,表明黄毛草莓、五叶草莓叶片蜡质对高温的响应较为相似。周玲艳等研究发现,40 ℃高温处理可造成水稻叶片角质层蜡质结构的改变和蜡质覆盖减少,导致非气孔性失水增加,这种响应可能既是一种保护,也是一种伤害[30]。
综上所述,黄毛草莓的耐热性要高于五叶草莓,40 ℃高温处理对黄毛草莓的有害效应要低于五叶草莓。黄毛草莓与五叶草莓对高温的响应策略有所不同,黄毛草莓可能通过增加角质层厚度、降低气孔密度和蒸腾速率来减少高温所引起的水分丢失,通过将更多物质分配到地上部分、提高超氧化物歧化酶活性来提高植株抗高温胁迫的能力;而五叶草莓可能通过增加气孔密度、提高气孔开度、增加蒸腾速率来降低高温对植株的伤害,并通过将更多的物质分配到地下部分来增强植株对高温胁迫的耐受能力。野生黄毛草莓具有的较高耐热性及抗热能力可用作我国草莓品种改良的特异种质资源,对改善我国草莓品种遗传背景简单、抗逆、抗病性差等缺点具有重要的价值。
:
[1]Christou A,Filippou P,Manganaris G A,et al. Sodium hydrosulfide induces systemic thermotolerance to strawberry plants through transcriptional regulation of heat shock proteins and aquaporin[J]. BMC Plant Biology,2014,14(1):42-52.
[2]Kadir S,Sidhu G,Al-Khatib K. Strawberry (FragariaxananassaDuch.) growth and productivity as affected by temperature[J]. HortScience,2006,41(6):1423-1430.
[3]郑 毅. 温度胁迫对草莓叶片光合作用的影响[D]. 合肥:安徽农业大学,2005:9-10.
[4]钟霈霖,乔 荣,王天文. 温度对夏秋草莓可溶性固形物含量的影响[J]. 贵州农业科学,2006,34(增刊1):53-54.
[5]朱 薇,杨明挚. 中国野生草莓资源研究及利用进展[J]. 中国南方果树,2014,41(4):50-58.
[6]李洪雯,刘建军,陈克玲,等. 四川及其周边地区野生草莓资源调查、收集与评价[J]. 植物遗传资源学报,2012,13(6):946-951.
[7]张常隆,李扬苹,冯玉龙,等. 表型可塑性和局域适应在紫茎泽兰入侵不同海拔生境中的作用[J]. 生态学报,2009,29(4):1940-1946.
[8]莫惠栋. Logistic方程及其应用[J]. 江苏农学院学报,1983,4(2):53-57.
[9]Lafta A M,Lorenzen J H. Effect of high temperature on plant growth and carbohydrate metabolism in potato[J]. Plant Physiology,1995,109(2):637-643.
[10]Marcum K B. Cell membrane thermostability and whole-plant heat tolerance ofKentuckybluegrass[J]. Crop Science,1998,38(5):1214-1218.
[11]Bewley J D. Physiological aspects of desiccation tolerance[J]. Annual Review of Plant Biology,1979,30(1):195-238.
[12]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:260-261.
[13]周小云,陈信波,徐向丽,等. 稻叶表皮蜡质提取方法及含量的比较[J]. 湖南农业大学学报,2007,33(3):273-276.
[14]夏莹莹,叶 航,马锦林,等. 4个油茶品种的半致死温度与耐热性研究[J]. 中国农学通报,2012,28(4):58-61.
[15]孙 震. 十种野生地被植物生态适应性的研究[D]. 北京:北京林业大学,2006:16-17.
[16]杜国栋,吕德国,赵 玲,等. 高温对仁用杏光合特性及PSⅡ光化学活性的影响[J]. 应用生态学报,2011,22(3):701-706.
[17]Luomala E M,Laitinen K,Sutinen S,et al. Stomatal density,anatomy and nutrient concentrations of Scots pine needles are affected by elevated CO2and temperature[J]. Plant Cell and Environment,2005,28(6):733-749.
[18]李海波,李全英,陈温福,等. 氮素不同用量对水稻叶片气孔密度及有关生理性状的影响[J]. 沈阳农业大学学报,2003,34(5):340-343.
[19]尚永申.水稻耐高温突变体hst(heat-shock tolerance)分子生理研究[D]. 杭州:浙江大学,2011:20-21.
[20]Feierabend J,Mikus M. Occurrence of a high temperature sensitivity of chloroplast ribosome formation in several higher plants[J]. Plant Physiology,1977,59(5):863-867.
[21]Mohanty S,Grimm B,Tripathy B C. Light and dark modulation of chlorophyll biosynthetic genes in response to temperature[J]. Planta,2006,224(3):692-699.
[22]陈 芳,郑炜君,李盼松,等. 小麦耐热性鉴定方法及热胁迫应答机理研究进展[J]. 植物遗传资源学报,2013,14(6):1213-1220.
[23]Hutchings M J. Foraging in plants:the role of morphological plasticity in resource acquisition[J]. Advances in Ecological Research,1994,25(3):159-238.
[24]金不换,陈雅君,吴艳华,等. 早熟禾不同品种根系分布及生物量分配对干旱胁迫的响应[J]. 草地学报,2009,17(6):813-816.
[25]Vile D,Pervent M,Belluau M,et al. Arabidopsis growth under prolonged high temperature and water deficit:independent or interactive effects?[J]. Plant Cell and Environment,2012,35(4):702-718.
[26]吕晓敏,王玉辉,周广胜,等. 温度与降水协同作用对短花针茅生物量及其分配的影响[J]. 生态学报,2015,35(3):753-760.
[27]Wahid A,Gelani S,Ashraf M,et al. Heat tolerance in plants:an overview[J]. Environmental and Experimental Botany,2007,61(3):199-223.
[28]Edelmann H G,Neinhuis C,Bargel H. Influence of hydration and temperature on the rheological properties of plant cuticles and their impact on plant organ integrity[J]. Journal of Plant Growth Regulation,2005,24(2):116-126.
[29]王家训. 柑桔叶片不同叶龄角质层和蜡质的季节变化[J]. 华中师范大学学报:自然科学版,1989,23(2):247-252.
[30]周玲艳,姜大刚,李 静,等. 逆境处理下水稻叶角质层蜡质积累及其与蜡质合成相关基因OsGL1表达的关系[J]. 作物学报,2012,38(36):1115-1120.
