宁夏罗山步甲昆虫群落多样性研究
2018-05-24秦伟春田会刚王新谱杨贵军
秦伟春,田会刚,王新谱,杨贵军
(1宁夏罗山国家级自然保护区管理局,宁夏 红寺堡 751900;2宁夏大学a农学院,b生命科学学院,宁夏 银川750021)
步甲科(Carabidae)昆虫是栖息于地表数量最庞大、种类最丰富的昆虫类群之一,不同生态区域植被生境类型的步甲昆虫多样性已成为生物多样性的重点研究内容[1-5],其物种的组成及数量变化可作为环境变化的重要监测指标[6-7]。已有研究表明,植物的种类组成和数量与步甲的种类及其种群结构密切相关[8-9],凋落物层的种类和数量变化影响步甲群落结构[10],不同土地利用方式对环境产生干扰从而也影响步甲多样性[11]。
罗山作为宁夏三大天然林区之一,其地理位置独特,生态功能极其重要,该地区的昆虫研究逐步受到重视,但目前主要侧重于昆虫的分类和区系研究[12],而关于昆虫栖息地环境因素对昆虫群落组成和多样性变化方面的影响涉及较少。本研究以罗山中段作为研究区,以对环境比较敏感的步甲昆虫为研究对象,调查不同生境步甲的群落组成和多样性,以期探讨该地区步甲昆虫分布和物种多样性与生境的关系,掌握该地区物种多样性变化规律,为该地区生物多样性的长期监测和保护提供依据。
1 材料与方法
1.1 研究区概况与样地设置
研究区域位于宁夏罗山国家级自然保护区境内,地理位置在宁夏中部同心县和红寺堡区境内(37°11′~37°25′N,106°04′~106°24′E),地处鄂尔多斯台地西部与黄土高原北部的衔接地带,区域高度1 560~2 624.5 m,山势挺拔,山体四周地势平缓,为荒漠植被,是宁夏中部干旱带重要的天然水源涵养和生态屏障区,素有旱海明珠、荒漠翡翠之美誉。该区域属中温带干旱大陆性气候,年平均气温8.8 ℃,年平均降水量261.8 mm,全年冷暖干湿四季分明,且冬春长、夏秋短,大风、沙暴、干旱热干风、霜冻、冰雹等灾害性天气较多。土壤类型主要有灰褐土、灰钙土、粗骨土。受地势影响,植被分布具有较明显的垂直分布特征,分为亚高山草甸、针叶林、针阔混交林、落叶灌丛、山前荒漠半荒漠草原等[12]。根据该区域内生境类型和海拔高度设置了8个样地。
亚高山草甸样地A(Subalpine meadow A,SMA):位于罗山山顶(红庙沟沟顶山脊)地势低洼处,水分条件相对较好,植被种类繁多,植被盖度大于90%,海拔2 560 m,属于典型的亚高山草甸植被类型。典型植被有秦艽(Gentianamacrophylla)、珠芽蓼(Polygonumviviparum)、垂穗披碱草(Elymusnutans)、线叶柴胡(Bupleurumangustissimum)、鹅绒委陵菜(Potentillaanserine)、铁杆蒿(Tripoliumvulgare)、小花风毛菊(Saussureaparviflora)等。
亚高山草甸样地B(Subalpine meadow B,SMB):位于罗山山脊处阳坡地带,为缓坡地形,水分条件、土壤条件较差,土壤中存在少量石质,海拔2 547 m,属亚高山针茅草甸。典型植被有大针茅(Stipagrandis)、扁穗冰草(Agropyroncristatum)大黄(Rheumpalmatum)、中国马先蒿(Pedicularischinensis)等。
针叶林样地A(Coniferous forests A,CFA):位于东风沟沟脑,海拔2 499 m,为青海云杉(Piceacrassifolia)纯林。林冠层较大,郁闭度较高(在0.6以上),林中常伴有山杨(Populusdavidiana)、油松(Pinustabuliformis),大部分林下盖度不足10%,部分林荫处地表覆有耐荫喜湿的苔藓,盖度可达90%。
针叶林样地B(Coniferous forests B,CFB):位于东风沟的油松密林处,海拔2 299 m,为油松(Pinustabuliformis)纯林。土壤为灰褐土,较青海云杉林土壤干燥,多砾石,有机质厚25~40 cm,结构良好。林内郁闭度不大。林下有虎榛子(Ostryopsisdavidiana)、栒子(Cotoneasterorizontalis)、小叶忍冬(Loniceramicrophylla)等散生灌木。
针阔混交林样地(Mixed broadleaf-conifer forest,MBCF):位于东风沟油松林与阔叶林白桦(Betulaplatyphylla)混交林处,海拔2 270 m。郁闭度较小,林内透光较好。林下有华北紫丁香(Syzygiumaromaticum)、虎榛子、小叶忍冬、绣线菊(Spiraeasalicifolia)、栒子等散生。
阔叶林缘样地(Broad-leaved forest edge,BLFE):位于东风沟白桦林林缘地带,为阔叶林类型,海拔2 190 m,林草植被盖度达90%以上,林缘有华北紫丁香、栒子、大针茅(Stipagrandis)、北柴胡(Bupleurumchinense)等。
灌丛样地(Deciduous shrub,DS):位于东风沟灌木林地带,海拔2 138 m,群落盖度在75%以上,典型植被有灌木华北紫丁香、小叶忍冬,与虎榛子等混生,灌丛高1 m以上。草本植物主要有小红菊(Dendranthemachanetii)、火绒草(Leontopodiumalpinum)、苔草(Carextristachya)等中旱生或旱生草原植物。
荒漠半荒漠草地样地(Desert semi-desert grasslands,DSDG):位于东风沟沟口的洪积扇坡地上,为荒漠半荒漠草地,海拔1 825 m。主要分布有小半灌木猫头刺(Oxytropisaciphylla)、达乌里胡枝子(Lespedezadavurica)、刺旋花(Convolvulustragacanthoides)、长芒草(Stipabungeana)、糙隐子草(Cleistogenessquarrosa)等旱生和中旱生植物。
1.2 调查方法
采用巴氏罐诱法[13],以一次性塑料水杯(高9 cm,口径7.5 cm)作为陷阱容器,在距水杯口1.5~2.0 cm 处打几个小孔,避免降雨使杯子水满造成标本流失,埋入地下使杯口基本与地面保持水平。每个样地内设置10 m×10 m小样方,重复3次,间距不小于50 m,共设定24个。每个样方内以间距2 m棋盘式设置诱杯,共25个。引诱剂由醋、糖、工业酒精和水按2∶1∶1∶20的质量比混合,每个诱杯内放引诱剂30~40 mL,陷阱杯放置时间为3 d。在2015年5-8月每月中旬收集1次,步甲标本保存在体积分数75%酒精中,带回实验室制成针插标本,进行种类鉴定[12]和统计分析。
1.3 数据处理
优势种类、常见种类和稀有种类的划分标准为:个体数量>10%者为优势种类,个体数量≥1%~≤10%为常见种类,个体数量<1%者为稀有种类[3]。
分析评价的指标有4个:(1)物种丰富度(S)。为表征群落中包含物种数量的量度,一般用物种数表示。(2)Shannon-Wiener多样性指数(H′)。H′=-∑PilnPi,其中Pi=Ni/N,Pi为第i种个体数量占总个体数量的比率,Ni为第i种的个体数量,N为总个体数量;(3)均匀性指数(J)。J=H′/lnS,H′为Shannon-Wiener多样性指数,S为物种数;(4)优势度指数(D)。采用Berger-Parker指数表示,D=Nmax/N,其中Nmax为优势种的种群数量[14]。
选用SPSS 19.0中的单因素方差分析(One-way ANOVA)对步甲群落多样性特征进行差异显著性检验。采用CANOCO 5.0软件对步甲群落结构进行主成分分析(Principal component analysis, PCA)、排序并作图,排序数据进行ln(x+1)转化。
2 结果与分析
2.1 步甲昆虫群落物种组成
2.1.1 步甲群落的组成与个体数量 调查统计结果表明,8个样地共采集步甲昆虫3 749只,隶属于12属24种。不同生境类型步甲组成、个体数量及所占比例见表1。从物种组成上看,婪步甲属(Harpalus)有8种,占总种数的33.33%;步甲属(Carabus)和猛步甲属(Cymindis)各3种,分别占总种数的12.50%;伪葬步甲属(Pseudotaphoxenus)有2种,占总种数的8.33%;其余8种为单属、单种,占总种数的33.33%。
从个体数量来看,直角通缘步甲和弗氏步甲个体数量较多,分别占总个体数量的39.11%和20.42%,为研究区域的优势种类。短翅伪葬步甲、小细胫步甲、考氏肉步甲、锥步甲、刻步甲、皮步甲、肩猛步甲、直角婪步甲、谷婪步甲、大卫婪步甲、喜婪步甲、棒婪步甲、喜湿步甲和蒙古伪葬步甲等个体数量分别占总个体数量的1.01%~9.41%,为本地常见种类,占总种数的58.33%。双斑猛步甲、半猛步甲、赤胸长步甲、径婪步甲、广胸婪步甲、黄鞘婪步甲、锯步甲和单齿蝼步甲个体数量占总个体数量的比例均小于1%,为本地稀有种类,共8种,占总种数的33.33%。

表1 宁夏罗山不同生境类型样地步甲种类个体数量及所占比例Table 1 Number of individuals and proportion of Carabidae species in different habitats of Ningxia Luo Mountain
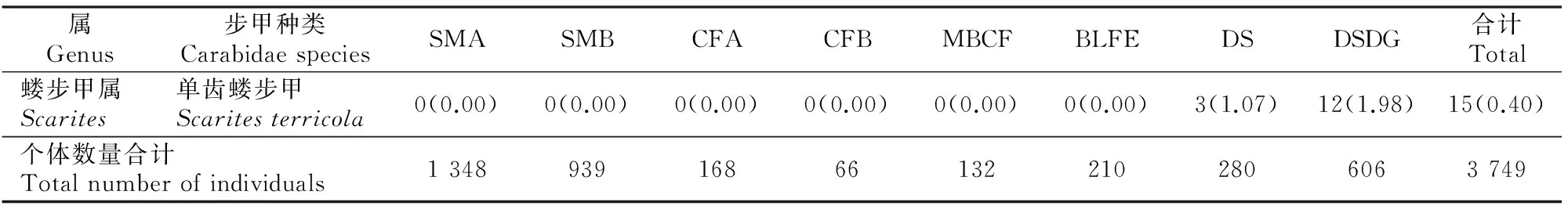
表1(续) Continued table 1
注:表中括号外数据为步甲个体数,括号内为其所占比例。
Note:The data out of parentheses is number of individuals and in parentheses is the percentage of individuals to total.
2.1.2 不同生境步甲的分布特点 从表1可以看出,不同生境步甲群落组成不同,样地SMA和SMB的优势种类为弗氏步甲和直角通缘步甲,样地CFA的优势种类为小细胫步甲、弗氏步甲、喜湿步甲和锯步甲,样地CFB的优势种类为小细胫步甲、喜湿步甲、锯步甲和直角通缘步甲,样地MBCF和BLFE的优势种类为直角通缘步甲、弗氏步甲和考氏肉步甲,样地DS的优势种类为短翅伪葬步甲、直角通缘步甲和大卫婪步甲,样地DSDG的优势种类为短翅伪葬步甲和直角通缘步甲。
从物种的分布来看,弗氏步甲、直角通缘步甲、考氏肉步甲、棒婪步甲和蒙古伪葬步甲等种类分布于6种以上的生境中,属于广谱分布种,其中直角通缘步甲在各生境均有分布,且在亚高山草甸生境分布数量最多。弗氏步甲仅在CFB生境无分布,也以亚高山草甸生境分布数量最多。考氏肉步甲仅在亚高山草甸无分布,其分布偏好于有林生境。蒙古伪葬步甲在针叶林以外的生境均有分布。棒婪步甲仅在油松林生境和荒漠半荒漠草地生境中无分布。小细胫步甲、喜湿步甲和锯步甲多在针叶林生境聚集,锥步甲、肩猛步甲和谷婪步甲多在光照充足的亚高山草甸和荒漠半荒漠草地中聚集。黄鞘婪步甲只分布在亚高山草甸生境中,单齿蝼步甲只分布在灌丛及其边缘相邻的荒漠草地生境中。
2.2 不同生境步甲分布的季节变化
宁夏罗山不同生境样地步甲昆虫的物种数、个体数量及优势种类的季节变化如图1所示。

图1 宁夏罗山不同生境中步甲物种和数量的季节变化Fig.1 Seasonal variation of species number and individuals of carabid beetles in different habitat plots of Ningxia Luo Mountain
从图1-A、B可以看出,亚高山草甸生境样地SMA和SMB在6-8月步甲物种数量都较高,针叶林生境样地CFA和CFB 7月步甲物种数量最高,针阔混交林样地MBCF和灌丛样地DS步甲物种数的高峰期均在6-7月,阔叶林缘样地BLFE的物种高峰期在7月,荒漠半荒漠样地DSDG的物种数最大值也出现在7月。样地SMA步甲个体数量在6和8月出现2次高峰,样地SMB则以6月份最高,其他生境样地步甲个体数量基本上随时间递增,针叶林样地CFA、CFB和荒漠半荒漠样地DSDG在8月份最高,样地MBCF、BLFE和DS的最大值出现在7月份。
由图1-C、D可以看出,优势种类直角通缘步甲在样地SMA和SMB数量最多,且在7月份有最大值,而在样地CFA中仅于6月份可采集到,在样地CFB中以7月份数量最多,在样地MBCF、BLFE和DS的最大值均出现在6月份,在样地DSDG中以5月份数量最多。弗氏步甲在油松林样地CFB无分布,在样地SMA、SMB和DSDG的个体数量最大值均出现在8月,在样地CFA、MBCF、BLFE和DS数量高峰值均出现在7月份。
2.3 步甲昆虫群落的多样性
从图2可以看出,不同样地的生境类型影响了步甲的物种数和个体数量,且物种种类和个体数量相关不显著,分布趋势不一致。从物种数方面分析,样地SMA和DS的步甲物种数最多,其次是样地BLFE和DSDG,再次是样地MBCF、SMB和CFA,最少的是样地CFB。从个体数量来看,不同样地物种个体数量由多到少依次表现为样地SMA>SMB>DSDG>DS>BLFE>CFA>MBCF>CFB。由此可见,在光照相对充足、草本覆盖度相对较高的亚高山草甸和荒漠半荒漠草地样地,步甲个体数量较多,但物种种类数量并不随上述两个自然生境条件下的个体数量多少呈线性分布。
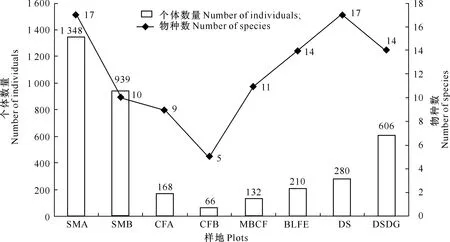
图2 宁夏罗山不同生境类型样地的步甲物种数和个体数量Fig.2 Number of individuals and carabid species in different habitat plots of Ningxia Luo Mountain
不同生境样地步甲群落多样性的分析结果见图3。由图3可知,不同生境样地的物种丰富度不同(F=5.491,P=0.000 7),其中灌丛样地DS物种数最多,亚高山草甸样地SMA次之,油松林样地CFB物种数最少。Shannon-Wiener多样性指数(H′)在不同生境间有极显著差异(F=3.876,P=0.005 9),8种样地H′依次表现为:DS>BLFE>DSDG>MBCF>SMA>CFA>CFB>SMB。均匀度指数在不同生境间也有极显著差异(F=11.189,P<0.001),其中以针叶林样地CFB最高,CFA次之,亚高山草甸样地SMA和SMB较低。优势度指数的变化趋势与Shannon-Wiener多样性指数的变化呈极显著性负相关(r=0.943,P<0.01),优势度指数在不同生境间差异极显著(F=5.358,P=0.000 9),其中以亚高山草甸样地SMB最高,样地SMA次之,灌丛样地DS最低。
2.4 不同生境样地步甲群落的PCA分析
对8个样地24种步甲的多度进行PCA分析,物种和样方的二维排序图(图4)表明,第1和第2排序轴对群落变化的解释度达到78.46%,能很好地反映不同生境样地步甲物种的分布。从图4还可以看出,8个样地的步甲昆虫群落可以聚为三类,其中灌丛样地DS、阔叶林缘样地BLFE和针阔混交林样地MBCF的步甲群落聚为一类,针叶林样地CFA和样地CFB的步甲昆虫群落聚为一类,亚高山草甸样地SMA、SMB和荒漠半荒漠草地样地DSDG的步甲昆虫群落可以聚为一类。

图柱上标不同小写字母表示在P<0.05水平差异显著Different lowercase letters mean significant difference at P<0.05图3 宁夏罗山不同生境样地步甲昆虫群落的多样性Fig.3 Comparison of carabid community diversity indices among different habitats of Ningxia Luo Mountain

sp1.小细胫步甲;sp2.考氏肉步甲;sp3.弗氏步甲;sp4.锥步甲;sp5.刻步甲;sp6.皮步甲;sp7.双斑猛步甲;sp8.半猛步甲;sp9.肩猛步甲;sp10.赤胸长步甲;sp11.直角婪步甲;sp12.谷婪步甲;sp13.大卫婪步甲; sp14.喜婪步甲; sp15.径婪步甲; sp16.广胸婪步甲; sp17.棒婪步甲; sp18.黄鞘婪步甲; sp19.喜湿步甲; sp20.锯步甲;sp21.直角通缘步甲; sp22.蒙古伪葬步甲; sp23.短翅伪葬步甲;sp24.单齿蝼步甲sp1.Agonum nitidum;sp2.Broscus kozlovi;sp3.Carabus vladimirskyi;sp4.Carabus glyptopterus;sp5.Carabus kruberi;sp6.Corsyra fusula;sp7.Cymindis binotata;sp8.Cymindis daimio;sp9.Cymindis scapularis;sp10.Dolichus halensis;sp11.Harpalus corporosus;sp12.Harpalus calceatus;sp13.Harpalus davidi;sp14.Harpalus optabilis;sp15.Harpalus salinus;sp16.Harpalus amplicollis;sp17.Harpalus bungii;sp18.Harpalus pallidipennis;sp19.Notiophilus aquaticus;sp20.Pristosiaproxima;sp21.Pterostichus gebleri;sp22.Pseudotaphoxenus mongolicus;sp23.Pseudotaphoxenus brevipennis;sp24.Scarites terricola图4 宁夏罗山不同生境样地步甲昆虫的PCA二维排序图Fig.4 Ordination diagram of principal component analysis on carabid beetles species in different plots of Ningxia Luo Mountain
3 讨 论
3.1 步甲种类组成与生境的关系
植被类型、土壤环境等诸多因素均会影响昆虫的空间分布。从本研究不同样地步甲物种组成的分析结果看,分布在罗山国家级自然保护区垂直植被带的8个样地具有一定的代表性,反映出生境的梯度变化对步甲昆虫群落结构的明显影响。亚高山草甸样地SMA和灌丛样地DS植被异质性较高,步甲物种数最多,各有17种,占研究区域物种总数的70.83%。样地SMA水分条件相对较好,植被丰富,植被盖度大于90%,较高的生境异质性为步甲昆虫分布提供了空间,而且该样地捕获的步甲个体数量也最多。样地SMB的植被多样性要低于样地SMA,其原因是由于该样地大针茅密度大,尤其到8月份降水增多,大针茅生长旺盛且竞争力强,其余草本营养空间有限,因而造成植被多样性减少,不利于更多种类地表昆虫的聚集,导致其步甲物种种类较少。针叶林样地步甲种类最少,其原因是其林下植被单一,透光性差,尤其是油松林样地CFB的步甲种类和数量均最少,主要是由于该样地较云杉林样地CFA的水分和土壤条件更差,该结果与已有研究结果[9]不一致,即山地针叶林的物种数和个体数量都较低,说明该研究区域山地针叶林面积较小,步甲区系种类主要体现了旱生和中旱生特点。阔叶林缘样地BLFE采集到的步甲昆虫有14种,占研究区域物种总数的58.33%,其原因可能是植被生境的边缘效应有助于提高昆虫的多样性[4,15]。荒漠半荒漠草地样地DSDG聚集了更多适应旱生的步甲种类,体现了该研究区域步甲组成的主要特点。
不同步甲昆虫的生物学特征体现了其对不同生境的适应性。弗氏步甲、直角通缘步甲等种类分布于6种以上的生境,属于栖境广布种,但分布并不均匀。优势种类在不同生境内的季节变化趋势也体现了其与不同生境植被生长季节的相关性。小细胫步甲、喜湿步甲和锯步甲趋向分布于阴湿的针叶林环境,而蒙古伪葬步甲在针叶林没有分布。锥步甲、肩猛步甲和谷婪步甲趋向于在光照充足的亚高山草甸和荒漠半荒漠草地中分布。此外,PCA分析排序中的每个物种在多个环境梯度上的空间位置,进一步反映了其在分类学性状上的差异[16-17],步甲属(Carabus)的弗氏步甲、锥步甲和刻步甲位置靠近,伪葬步甲属(Pseudotaphoxenus)的蒙古伪葬步甲和短翅伪葬步甲位置也靠近,其生境选择上不偏好阴湿环境。
3.2 步甲群落多样性与生境的关系
宁夏罗山垂直植被带8个不同生境步甲昆虫群落的多样性,反映出环境因素对维持群落多样性的重要作用,生境的相似性在一定程度上决定着地表甲虫群落组成的相似性,聚类分析结果显示,生境相近的样地步甲昆虫群落结构也相似。生境异质性是维持生物多样性的主要因素,生境差异程度增加,相似性系数相应降低。灌丛样地DS的Shannon-Wiener多样性指数最高,一方面由于灌丛植被异质性较高,另一方面是灌丛“顶蓬”效应为昆虫提供了丰富的食物资源和相对温和安全的产卵场所[18]。针叶林生境样地的Shannon-Wiener多样性指数较低,尤其是针叶林样地CFB多样性指数最低。海拔对昆虫的分布格局呈现主导控制作用,本研究结果表明,步甲昆虫群落Shannon-Wiener多样性指数基本表现出随垂直带即海拔升高而降低的现象,这与很多研究结果[9,19]相一致。步甲物种丰富度的变化以中段海拔较低,该结果与普遍的单峰分布格局不一致[20],可能与中段海拔尤其是针叶林林下植被相对较为单一及步甲的生物学特性有关。而均匀性指数的变化趋势呈现为CFB>CFA>DS>MBCF>BLFE>DSDG>SMA>SMB,与Shannon-Wiener多样性变化趋势不一致,可能是因为常绿针叶林下地表环境相对稳定,地表步甲昆虫群落结构也较为稳定,亚高山草甸和荒漠半荒漠草地生境年四季气候变化、植被季节更替对地表影响较大,步甲群落结构在研究时间段均匀性指数较低,但并不能说明这2种生境的步甲群落结构稳定性低,群落的稳定性还体现在自身的恢复能力上,因此尚需进行多年季节演替规律的调查研究。
[参考文献]
[1] 贺 奇,王新谱,杨贵军.宁夏盐池荒漠草原步甲物种多样性 [J].生态学报,2011,31(4):923-932.
He Q,Wang X P,Yang G J.Species diversity of carabid beetles in desert-steppe in Yanchi of Ningxia,China [J].Acta Ecologica Sinica,2011,31(4):923-932.
[2] Jia Y Z,Zhao X H,Meng Q F.Composition and diversity of ground-dwelling beetle (Coleoptera) along a succession gradient in broad-leaved and Korean pine mixed forest in the Changbai Mountains, China [J].Chinese Journal of Applied & Environmental Biology,2011,17(6):797-802.
[3] 杨贵军,贾彦霞,王新谱,等.苜蓿-荒漠草地交错带步甲昆虫多样性 [J].环境昆虫学报,2015,37(3):483-491.
Yang G J,Jia Y X,Wang X P,et al.Diversity of carabid beetles community across alfalfa-desert grassland ecotone in Yanchi of Ningxia [J].Journal of Environmental Entomology,2015,37(3):483-491.
[4] 郭 瑞,王义平,翁东明,等.浙江清凉峰不同植物群落步甲物种多样性及其与环境因子的关系 [J].浙江农林大学学报,2016,33(4):551-557.
Guo R,Wang Y P,Weng D M,et al.Carabid beetle(Coleoptera:Carabidae)species diversity and environmental factors in biotopes of Zhejiang Qingliangfeng National Nature Reserve, China [J].Journal of Zhejiang A&F University,2016,33(4):551-557.
[5] 王 晶,吕昭智,殷 飞.干旱区景观异质性对地表甲虫多样性的影响 [J].环境昆虫学报,2016,38(1):67-76.
Wang J,Lü Z Z,Yin F.The influence of landscape heterogeneity on diversity of ground beetles in arid regions [J].Journal of Environmental Entomology,2016,38(1):67-76.
[6] Antvogel H,Bonn A.Environmental parameters and microspatial distribution of insects: a case study of carabids in an alluvial forest [J].Ecography,2001,24(4):470-482.
[7] Molnar T,Magura T,Tóthmérész B,et al.Ground beetles(Carabidae) and edge effect in oak-hornbeam forest and grassland transects [J].Eur J Soil Biol,2001,37(4):297-300.
[8] 于晓东,周红章,罗天宏.神农架保护区大步甲和蜗步甲属生境选择与物种多样性研究 [J].生物多样性,2001,9(3):214-221.
Yu X D,Zhou H Z,Luo T H.Habitat distribution and species diversity ofCarabusandCychrusin Shennongjia Nature Reserve,Hubei province [J].Biodiversity Science,2001,9(3):214-221.
[9] Tyler G.The ground beetle fauna(Coleoptera:Carabidae)of abandoned fields, as related to plant cover, previous management and succession stage [J].Biodiversity and Conservation,2008,17(1):155-172.
[10] Haila Y,Niemelä J.Leaf litter and the small-scale distribution of carabid beetles (Coleoptera: Carabidae) in the boreal forest [J].Ecography,1999,22(4):424-435.
[11] 王建芳,王新谱,李秀敏, 等.不同土地利用方式对河北坝上步甲物种多样性的影响 [J].昆虫学报,2010,53(10):1127-1134.
Wang J F,Wang X P,Li X M,et al.Influence of land uses on species diversity of carabid beetles (Coleopeta:Carabidae) in Bashang region,Hebei,Northern China [J].Acta Entomol Sin,2010,53(10):1127-1134.
[12] 杨贵军,王新谱,仇智虎.宁夏罗山昆虫 [M].银川:阳光出版社,2011:165-168.
Yang G J,Wang X P,Qiu Z H.Insect fanua of Luo Mountains in Ningxia [M].Yinchuan:Sunshine Publishing House,2011:165-168.
[13] Ulyshen M D,Hanula J L,Horn S.Using malaise traps to sample ground beetles (Coleoptera:Carabidae) [J].The Canadian Entomologist,2005,137(2):251-256.
[14] 马克平,刘玉明.生物多样性测度方法:Ⅰ.α-多样性(下) [J].生物多样性,1994,2(4):231-239.
Ma K P,Liu Y M.Measurement of biotic community diversity:Ⅰ.α diversity(Part 2) [J].Biodiversity Science,1994,2(4):231-239.
[15] 于晓东,罗天宏,周红章,等.边缘效应对卧龙自然保护区森林-草地群落交错带地表甲虫多样性的影响 [J].昆虫学报,2006,49(2):277-286.
Yu X D,Luo T H,Zhou H Z,et al.Influence of edge effect on diversity of ground-dwelling beetles across forest-grassland ecotone in Wolong Natural Reserve,Southwest China [J].Acta Entomologica Sinica,2006,49(2):277-286.
[16] Irmler U,Hoernes U.Assignment and evaluation of ground beetle (Coleoptera:Carabidae) assemblages to sites on different scales in a grassland landscape [J].Biodiversity and Conservation,2003,12(7):1405-1419.
[17] 杨贵军,贺 奇,王新谱.盐池四墩子拟步甲昆虫群落组成与环境因子的相关性 [J].应用生态学报,2010,21(9):2375- 2382.
Yang G J,He Q,Wang X P.Darkling beetle community structure and its relations with environmental factors in Sidunzi of Yanchi,Ningxia,China [J].Chinese Journal of Applied Ecology,2010,21(9):2375-2382.
[18] Liu J L,Li F R,Liu C A,et al.Influences of shrub vegetation on distribution and diversity of a ground beetle community in a Gobi desert ecosystem [J].Biodiversity and Conservation,2012,21(10):2601-2619.
[19] Ikeda H,Tsuchiya Y,Nagata N,et al.Altitudinal life-cycle and body-size variation in ground beetles of the genusCarabus(subgenusOhomopterus) in relation to the temperature conditions and prey earthworms [J].Pedobiologia,2012,55(2):67-73.
[20] Sanders N J,Rahbek C.The patterns and causes of elevational diversity gradients [J].Ecography,2012,35(1):1-3.
