甘肃亚高山云杉人工林下植物种群生态位特征
2018-05-23冯宜明陈学龙侯亚丽闫倩倩李社勤徐德才
冯宜明,陈学龙,齐 瑞,赵 阳,侯亚丽,闫倩倩,李社勤,徐德才
(1.甘肃省白龙江林业管理局林业科学研究所,甘肃 兰州 730070; 2.甘肃白龙江森林生态系统国家定位观测研究站,甘肃 武都 746000; 3.甘肃省白龙江林管局舟曲林业局,甘肃 舟曲 746300)
生态位是近代理论生态学的一个主要内容,其致力于探讨生物对环境的要求、环境对生物的影响及其相互作用规律[1-2]。自Grinnell将生态位引入生态学研究领域以来,国内外学者对生态位理论[3-4]、生态位特征[5-6]、生态位测度[7-8]进行了大量的研究,广泛应用于物种间关系[9]、群落结构及演替[10]和种群进化[11]等方面,成为物种共存、竞争以及种群在群落中所处地位的评价手段,同时,也是生态学中重要的基础理论和最为活跃的研究领域之一[12-13]。种群生态位的研究不仅可以定量反映物种与环境的相互关系、种群对资源的利用能力,还可反映群落稳定性[14]。因此,对多种植被类型生态位的研究已有诸多报道,如针阔叶林[15-17]、荒漠植被[18-20]、高寒草原[21-22]等。
云杉(Piceaasperata)是我国西北和西南地区具有代表性的森林树种,也是甘肃的主要造林树种,具有适应性强、生长快的特点[23]。目前国内外对云杉群落的研究主要集中在群落结构[24]、生物量和碳密度[25]、生产力[26]以及林下植物多样性[27]等方面。林下灌草层是森林生态系统的重要组成部分,维持着整个系统物质循环和能量交换以及结构和功能的稳定性,对林下灌、草植物种群生态位的研究,可以阐述种群的种间关系及利用环境资源的状况,揭示种群的动态变化以及在群落发展进程中的地位和和功能,预测灌、草群落演替,提高森林生态系统中气候、水肥等资源的优化利用率;同时对该区域群落生态位数据进行补充,可推动生态位研究领域的积极发展。因此,以白龙江流域云杉人工林下的灌木、草本层为研究对象,采用Levins、Shannon-Wiener 生态位宽度指数和Levins生态位重叠指数,对云杉人工林下主要种群的生态位进行定量分析,旨在揭示各种群对资源的利用状况和生态适应性,选择适合于该区域云杉人工林的物种结构及共存模式,从而为林下生物多样性保育和森林经营管理提供理论参考。
1 材料与方法
1.1 研究区概况
甘肃省白龙江林区沙滩林场(102°02′-104°22′ E,33°34′-33°46′ N),地处青藏高原东北边缘与秦岭西段交汇处,具有典型的高山地形。地势起伏大,高差悬殊,平均坡度为30°~35°。土壤以棕色灰化土、棕色森林土、褐色森林土为主。属高山内陆性气候,年降水量1 023.1 mm,年蒸发量918.8 mm,年均气温4.3 ℃,极端高温29.1 ℃,极端低温-28.5 ℃,无霜期100 d[28]。林场经营总面积3.1万hm2,活立木总蓄积量为207.8万m3,森林覆盖率为74.6%。主要乔木树种有云杉、岷江冷杉(Abiesfaxoniana)、青扦(Piceawilsonii)、油松(Pinustabulaeformis)、落叶松(Larixgmelini)等;主要灌木有针刺悬钩子(Rubuspungens)、箭竹(Sinarundinarianitida)、毛花忍冬(Loniceratrichosantha)、扁刺蔷薇(Rosasweginzowii)、锐齿臭樱(Maddeniaincisoserrata)、藤山柳(Clematoclethrascandens)等。该区90%的粗枝云杉人工林于1980年前后营造。
1.2 研究方法
1.2.1样地设置与调查 2013年7月在沙滩林场沿海拔梯度(2 300~2 900 m)垂直于河道设置具有代表性的云杉乔木样地22块,样地大小为20 m×20 m,样地间隔大于200 m,调查每块样地乔木的种类、株数、胸径、树高、冠幅等指标。并于8-9月在乔木样地中心及四角布设5个2 m×2 m灌木样方,共计110个,调查记录样方内灌木种类、数量、基径、高度、盖度等。在灌木样方的中心位置布设一个1 m×1 m的草本样方,共计110个,进行草本植物的种类、个体数目、高度、盖度、频度等调查。
1.2.2重要值计测
灌木的重要值计算公式:重要值=(相对密度+相对高度+相对盖度+相对优势度)/4;
草本的重要值计算公式[29]:重要值=(相对密度+相对盖度+相对频度+相对高度)/4;
结合样地内植物群落的记录情况和每个样方内植物种数的均值情况,参考重要值在灌木层和草本层各选取15个物种进行生态位研究。
1.2.3生态位宽度计测
生态位宽度采用Levins和Shannon-Wiener生态位宽度指数进行测度[30]。

1.2.4生态位重叠计测 采用Levins生态位重叠指数计测。
式中:Lih为物种i重叠物种h的生态位重叠指数,B(L)i为物种i的Levins生态位宽度指数;Lhi为物种h重叠物种i的生态位重叠指数,B(L)h为物种h的Levins生态位宽度指数[31]。
1.2.5种群生态响应的测定 采用以下公式计算。
R=B(L)/ΔLih(i=h)。
式中:ΔLih为i种群占用其他种群资源量与h种群被其他种群侵占的资源量之差值;R为生态响应速率,B(L)为Levins生态位宽度指数[32]。
2 结果与分析
2.1 生态位宽度
云杉人工林下主要种具有的Shannon-Wiener和Levins生态位宽度值的总体变化趋势较为一致(表1)。灌木层针刺悬钩子、箭竹、秀丽莓、红桦幼苗、毛花忍冬、岷江冷杉幼苗、扁刺蔷薇、锐齿臭樱、藤山柳9个宽度值较大种的排序相同,其B(SW)在1.869 7~2.685 0,B(L)在0.241 0~0.573 2,具有较强的资源利用能力,是灌木层的优势种(表1)。两种测算公式测度的生态位宽度值较小的3个种是陕甘花楸、川滇柳、勾儿茶,说明这3种灌木在当前群落中对资源的竞争利用能力较弱,地位不明显。草本层中东方草莓、鸭茅、紫花碎米荠具有较大的生态位宽度,B(SW)与B(L)分别为3.207 4、3.169 2、3.127 9和0.976 9、0.944 3、0.907 4(表1),是草本层的优势种,在所调查的全部样方中均有出现,分布较为广泛,适应环境能力强,因此生态位宽度较大。囊瓣芹、长籽柳叶菜、风毛菊的生态位宽度则较小,对环境资源有较强的选择性。
2.2 生态位重叠
云杉人工林下灌木中箭竹-藤山柳、针刺悬钩子-山梅花、秀丽莓-红脉忍冬、针刺悬钩子-毛花忍冬4个种对之间的生态位重叠较大,分别为0.038 9、0.036 1、0.034 5和0.034 3(表2);草本中东方草莓-风毛菊、鸭茅-囊瓣芹、鸭茅-掌叶橐吾、冷蕨-粗茎鳞毛蕨和鸭茅-风毛菊4个种对之间的生态位重叠较大,分别为0.041 8、0.041 2、0.040 9、0.040 8和、0.040 8(表3),生态位重叠较高的种对均出现在生态位宽度较大的种群与其他种群之间,表明各种对具有相似的生物学特征或对生境因子有较强的互补性需求。但也有生态位较窄的物种之间出现较高的生态位重叠,如灌木层红脉忍冬和勾儿茶之间的生态位重叠值为0.0298,位居210个种对的第7位;草本层高山露珠草和囊瓣芹之间的生态位重叠值为0.0296,而生态位宽度值分居15个种的第12和13位。
对于同一个种对而言,Lih较高,Lhi则不一定高,如灌木层针刺悬钩子-山梅花的重叠值为0.036 1,而山梅花-针刺悬钩子仅为0.014 7;草本层东方草莓-风毛菊的重叠值为0.041 8,而风毛菊-东方草莓仅为0.019 3。综合来看,粗枝云杉人工林下主要种群的生态位重叠程度较低,灌木层Lih变动范围为0~0.038 9,Lhi变动范围为0~0.036 1,而且有85%以上的种对集中在0~0.020 0(表2);草本层Lih变动范围为0~0.041 8,Lhi变动范围为0~0.040 9(表3),表明云杉人工林下主要物种之间对资源利用的相似性较低。
2.3 生态响应
在云杉人工林下的灌木层,发展能力较强的是红桦幼苗、秀丽莓、针刺悬钩子,箭竹发展能力最弱;岷江冷杉幼苗、毛花忍冬、扁刺蔷薇等属于衰退性物种(表4),在红桦幼苗、秀丽莓等不断进攻下,它们的生态空间有逐渐缩小的趋势,其中冷杉幼苗表现最明显,其次为毛花忍冬和扁刺蔷薇,主要是因为红桦幼苗、秀丽莓的生态位宽度大于冷杉幼苗和毛花忍冬,它们对环境的适应性和对资源的利用能力也比岷江冷杉幼苗等衰退性物种强,因此,在有限的环境资源范围内岷江冷杉幼苗的生态空间会越来越小。
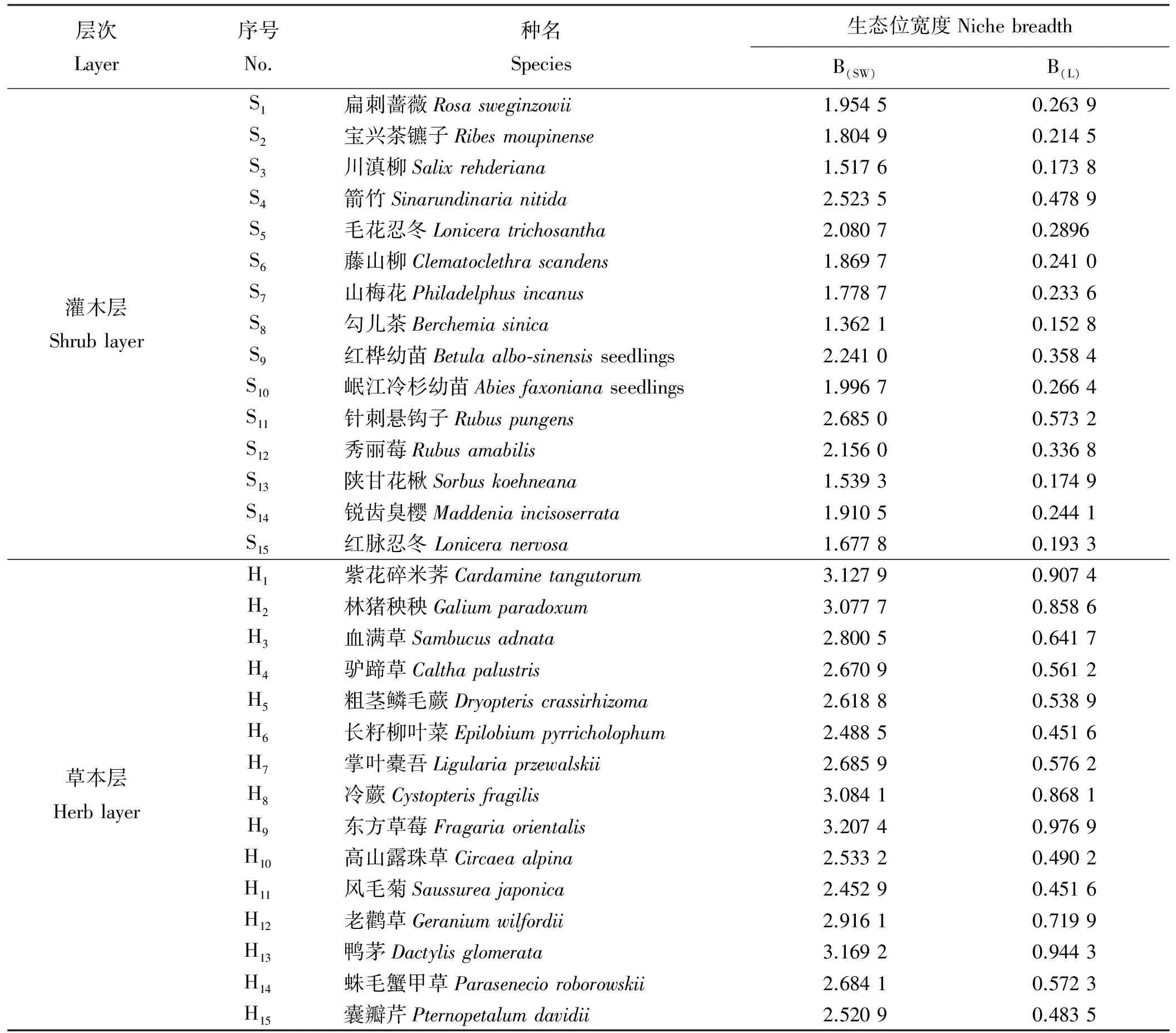
表1 云杉人工林下优势种生态位宽度值Table 1 Niche breadth of dominant species in the understory of a Picea asperata plantation
草本层发展能力较强的是老鹳草,远远超过其他种,东方草莓的发展能力最弱;血满草、掌叶橐吾、蛛毛蟹甲草和驴蹄草等属于衰退性物种(表5),在老鹳草、林猪秧秧等不断进攻下,它们的生态空间有逐渐缩小的趋势,其中血满草表现最明显,其次为掌叶橐吾和蛛毛蟹甲草。
3 讨论
生态位宽度揭示了物种对环境资源的利用状况,也表征了物种的生态适应性和分布幅度[33-34]。本研究应用Shannon-Wiener 和Levins生态位宽度公式对灌草层主要植物种生态位宽度测度的结果基本一致。灌木层中,针刺悬钩子、箭竹、秀丽莓具有较大的生态位宽度,这与它们的生物学特性密切相关,针刺悬钩子、箭竹、秀丽莓较耐阴,因而幼苗可在郁闭度较高的云杉林下繁殖更新,广泛分布。草本层中,东方草莓、鸭茅、紫花碎米荠具有较大的生态位宽度,这与其对环境的适应性和对资源的利用能力有关[35]。如东方草莓和鸭茅种群,在所有调查样方都有出现且数量多,对环境资源的利用比较充分,所以它们的生态位宽度较大。
生态位重叠作为衡量种间生态相似性的指标与种间竞争有着一定的联系[36]。具有相似环境要求的物种间生态位重叠较大,如囊瓣芹和掌叶橐吾都是比较耐阴的物种,它们与鸭茅都具有相近的生态特征并在生境因子上具有互补性,所以鸭茅-囊瓣芹、鸭茅-掌叶橐吾之间的生态位重叠值较高。总体看来,云杉人工林下植物种间的生态位重叠较低,物种对资源利用的相似性低,可能是因为随着乔木层郁闭程度的变化,林下植物也在不断的适应新环境,群落物种尚未达到稳定,王伟伟等[2]和钟宇等[8]对青海云杉(Spruceplantation)、巨桉(Eucalyptusgrandis)人工林草本层生态位的研究,得出了相似的结论。

表2 灌木层优势种生态位重叠值Table 2 Niche overlap of dominant species in the shrub layer
S1, S2, S3, …, S15所代表的物种与表1相同。
Species represented by S1, S2, S3, …, S15are same as Table 1.

表3 草本层优势种生态位重叠值Table 3 Niche overlap of dominant species in the herb layer
H1, H2, H3, …, H15所代表的物种与表1相同。
Species represented by H1, H2, H3, …, H15are same as Table 1.

表4 云杉人工林下优势灌木物种发展或衰退状况Table 4 Aggression or declining trends of the dominant shrub species in the understory of a Picea asperata plantation
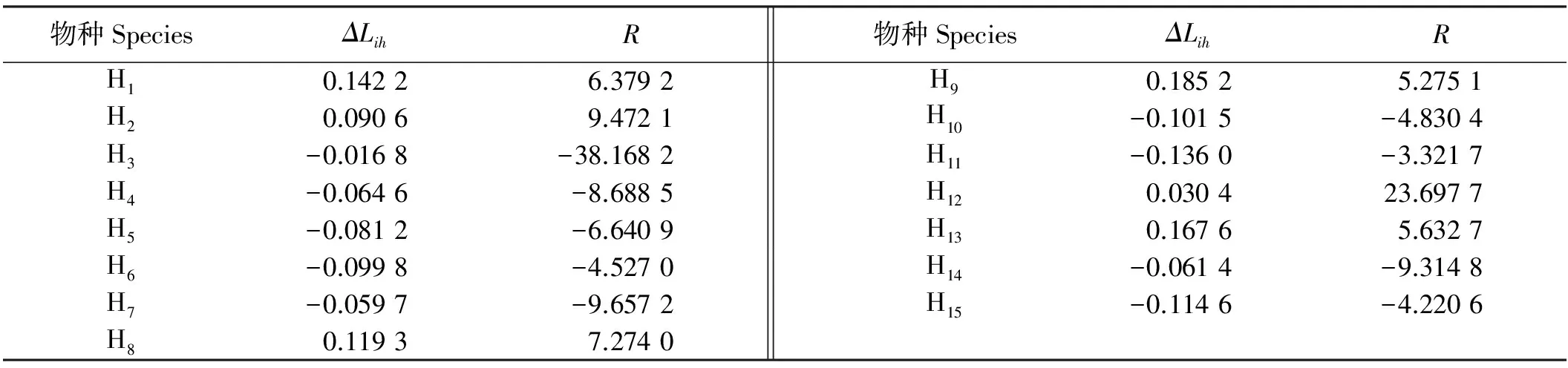
表5 云杉人工林下优势草本物种发展或衰退状况Table 5 Aggression or declining trends of the dominant herbaceous species in the understory of a Picea asperata plantation
生态位宽度和生态位重叠是生态位测度的两个方面,二者既联系又有区别。一般认为,生态位宽度大的种群对资源的利用能力较强,分布范围广,因而与其他种群生态位重叠的几率高[37]。然而本研究发现,生态位宽度较大的种群之间出现了较小的生态位重叠,这是因为生态位宽度反映的是一个种群在资源序列上占据空间的能力,而生态位重叠考虑的则是种群之间的竞争和共存关系,是建立在物种生物学特性和生态因子需求基础上的,因此,具有较大生态位宽度种群之间的生态学特性不同,对资源段位要求不完全一致,如若环境中共用的资源较为丰富,就会导致生态位重叠不高[38];生态位宽度较小的灌木红脉忍冬和勾儿茶、草本高山露珠草和囊瓣芹两个种对间却出现了极高的生态位重叠,这主要是由于红脉忍冬和勾儿茶,高山露珠草和囊瓣芹之间的生物学特性很相似,对环境的要求相同所致。这也反映出生态位重叠与生态位宽度之间的关系较为复杂,并没有绝对的正相关关系,这与柴宗政等[39]、胡正华等[40]、陈艳瑞和尹林克[41]的研究结论一致。
从生态响应的角度来看,在灌木层植物中发展性最强的是红桦幼苗,发展性最弱的是箭竹,冷杉幼苗衰退最为严重,这与它们对环境的适应性以及对资源的利用能力有关,红桦幼苗的生态位宽度大于冷杉幼苗,在有限的资源和日益激烈的竞争中,岷江冷杉幼苗的生态空间会越来越小。在草本层植物中发展性最强的是老鹳草,而东方草莓和鸭茅等生态位较宽的优势种发展最弱,血满草衰退最严重,这是由于生态响应是生物因子和非生物因子综合作用的结果,东方草莓、鸭茅生态位宽度较大,分布范围达到了峰值,随着环境条件的变化,它们适应新环境的能力有所下降,所以发展性减弱,而血满草这种较喜光照的植物在林下光照条件不足的环境中表现出明显的衰退趋势。按照这样的趋势发展演替,生态位较窄的衰退种将会逐渐消失,为了将它留存下来以提高物种多样性,应该通过人为干扰或改变环境资源状态的办法让优势种群释放一定的资源空间,从而增加它们生态位宽度。
本研究应用生态位理论,对云杉人工林下主要植物种群进行了一维资源位的生态位特征分析,清楚地了解了主要植物种群生态幅度和对资源利用的差异,探讨了优势种群在群落中的功能地位。但未引入基于物种营养生态位以及其他环境因子进行多维生态位的研究,还需在今后的研究中做进一步的探讨。
4 结论
1)云杉人工林下灌木层中,针刺悬钩子、箭竹、秀丽莓的生态位宽度较大,B(SW)和B(L)分别为2.685 0、2.523 5、2.156 0和0.573 2、0.478 9、0.336 8;草本层中,东方草莓、鸭茅、紫花碎米荠的生态位宽度较大,B(SW)和B(L)分别为3.207 4、3.169 2、3.127 9和0.976 9、0.944 3、0.907 4。它们对环境适应能力强,分布范围广,倾向于泛化种。2)云杉人工林下物种间生态位重叠程度整体较小。灌木箭竹与藤山柳、针刺悬钩子与山梅花、秀丽莓与红脉忍冬之间生态位重叠相对较大,草本东方草莓与凤毛菊、鸭茅与囊瓣芹、鸭茅与掌叶橐吾、冷蕨与粗茎鳞毛蕨之间也有相对较大的生态位重叠。生态位宽的种群可能产生较小的生态位重叠,生态位较窄的种群间也会产生较大的生态位重叠,生态位宽度与生态位重叠程度并没有绝对的正相关关系。3)从生态响应来看,灌木层的红桦幼苗和草本层的老鹳草发展性最强,今后可能会发展为云杉人工林下的优势种群,冷杉幼苗和血满草衰退最严重,生态空间逐渐缩小。
参考文献References:
[1] Leibold M A.The niche concept revisited mechanistic model and community context.Ecology,1995,76(5):1371-1382.
[2] 王伟伟,杨海龙,贺康宁,胡兴波,王辉,董梅.祁连山青海云杉人工林草本层生态位及生态响应研究.草地学报,2012,20(4):626-630.
Wang W W,Yang H L,He K N,Hu X B,Wang H,Dong M.Niche and ecological response of herb layer inSpruceplantationof Qilian Mountains.Acta Agrectir Sinica,2012,20(4):626-630.(in Chinese)
[3] Mueller L D,Altenberg L.Statistical inference on measures of niche overlap.Ecology,1985,66(4):1204-1210.
[4] 彭文俊,王晓鸣.生态位概念和内涵的发展及其在生态学中的定位.应用生态学报,2016,27(1):327-334.
Peng W J,Wang X M.Concept and connotation development of niche and its ecological orientation.Chinese Journal of Applied Ecology,2016,27(1):327-334.(in Chinese)
[5] Westman W E.Measuring realized niche spaces:Climatic response of chaparral and coastal sage scrub.Ecology,1991,72(5):1678-1684 .
[6] 刘金福,洪伟.格氏栲群落生态学研究:格氏栲林主要种群生态位的研究.生态学报,1999,19(3):347-352.
Liu J F,Hong W.A study on the community ecology ofCastanopsiskawakamii:Study on the niche of the main tree population inCastanopsiskawakamiicommunity.Acta Ecologica Sinica,1999,19(3):347-352.(in Chinese)
[7] 王刚,赵松岭,张鹏云,陈庆诚.关于生态位定义的探讨及生态位重叠计测公式改进的研究.生态学报,1984,4(2):119-127.
Wang G,Zhao S L,Zhang P Y,Chen Q C.On the definition of niche and the improved formula for measuring niche overlap.Acta Ecologica Sinica,1984,4(2):119-127.(in Chinese)
[8] 钟宇,张健,刘泉波,杨万勤,吴福忠,冯茂松.巨桉人工林草本层主要种群的生态位分析.草业学报,2010,19(4):16-21.
Zhong Y,Zhang J,Liu Q B,Yang W Q,Wu F Z,Feng M S.Niche characteristics of main herbage populations inEucalyptusgrandisplantation.Acta Prataculturae Sinica,2010,19(4):16-21.(in Chinese)
[9] 冉圣宏,金建君,薛纪渝.脆弱生态区评价的理论与方法.自然资源学报,2002,17(1):117-122.
Ran S H,Jin J J,Xue J Y.Assessment of vulnerable ecological regions:Theory and methodology.Journal of Natural Resource,2002,17(1):117-122.(in Chinese)
[10] Rosenthal G.Selecting target species to evaluate the success of wet grassland restoration.Agriculture Ecosystem & Environment,2003,98(3):227-246.
[11] 刘加珍,陈亚宁,张元明.塔里木河中游植物种群在四种环境梯度上的生态位特征.应用生态学报,2004,15(4):549-555.
Liu J Z,Chen Y N,Zhang Y M.Niche characteristics of plants on four environmental gradients in middle reaches of Tarim River.Chinese Journal of Applied Ecology,2004,15(4):549-555.
[12] Brathen K A,Ravolainen V T.Niche construction by growth forms is as strong a predictor of species diversity as environmental gradients.Journal of Ecology,2015,103:701-713.
[13] Fort H.Community diversity and total abundance:Quantitative predictions from competition niche theory.Ecological Complexity,2015,21:120-127.
[14] 林勇,艾训儒,姚兰,黄伟,陈斯.木林子自然保护区不同群落类型主要优势种群的生态位研究.自然资源学报,2017,32(2):223-234.
Lin Y,Ai X R,Yao L,Huang W,Chen S.Niches of main dominant species of different community types in Mulinzi Nature Reserve.Journal of Natural Resources,2017,32(2):223-234.(in Chinese)
[15] 潘高,张合平,潘登.南方红壤丘陵区3种森林群落内主要草本植物种群生态位特征.草业科学,2015,32(12):2094-2106.
Pan G,Zhang H P,Pan D.Niche characteristics of herb populations within three forest types in hilly red soil region of southern China.Pratacultural Science,2015,32(12):2094-2106.(in Chinese)
[16] 黄耀,王乃江,党鹏,裴乔.黄土高原沟壑区油松人工林林下主要种群生态位研究.内蒙古农业大学学报(自然科学版),2016,37(4):46-51.
Huang Y,Wang N J,Dang P,Pei Q.The study of the niche of the main plant population under region ofPinustabuliformisplantation in gully region of loess platelau.Journal of Inner Mongolia Agricultural University (Natural Science Edition),2016,37(4):46-51.(in Chinese)
[17] 潘高,张合平,潘登.湖南青羊湖国家森林公园针阔混交林主要种群生态位特征.中南林业科技大学学报,2017,37(2):69-75.
Pan G,Zhang H P,Pan D.Niche characteristics of dominant populations within mixed broadleaf-conifer forest in Qingyanghu National Park.Journal of Central South Forestry University & Technology,2017,37(2):69-75.(in Chinese)
[18] 韩路,王家强,王海珍,牛建龙,于军.塔里木荒漠绿洲过渡带主要种群生态位与空间格局分析.植物科学学报,2016,34(3):352-360.
Han L,Wang J Q,Wang H Z,Niu J L,Yu J.Niche and spatial distribution pattern analysis of the main populations of the Tarim desert-oasis ecotone.Plant Science Journal,2016,34(3):352-360.(in Chinese)
[19] 梁娜,靳瑰丽,安沙舟,赛米拉克孜·台外库力,迪丽努尔·玉苏甫江.伊犁绢蒿退化荒漠草地植物种群生态位的变化特征.草业科学,2013,30(12),2059-2065.
Liang N,Jin G L,An S Z,Saimilakezi·Taiwaikuli,Dilinuer·Yusupujiang.Plant population niche characteristics research of desertSeriphdiumtransiliensedegradation grassland.Pratacultural Science,2013,30(12):2059-2065.(in Chinese)
[20] 庞吉林,张克斌,王海星,王黎黎,王晓,苏朋飞.盐池县人工封育草地植物生态位动态研究.草业科学,2012,29(8):1285-1293.Pang J L,Zhang K B,Wang H X,Wang L L,Wang X,Su P F.Vegetable niche dynamic study of the enclosed desertification rangeland in Yanchi County.Pratacultural Science,2012,29(8):1285-1293.(in Chinese)
[21] 张爱宁,安沙舟,张蕊思,夏小伟,卡斯达尔·努尔旦别克,董磊.不同退化梯度下大尤尔都斯高寒草原植物种群生态位特征.草业科学,2017,34(2):302-309.
Zhang A N,An S Z,Zhang R S,Xia X W,Kasidaer·Nuerdanbieke,Dong L.Study on niche characteristics of the plant community in the alpine grassland of Big Youerdusi under a degeneration gradient.Pratacultural Science,2017,34(2):302-309.(in Chinese)
[22] 郑伟,董全民,李世雄,刘玉.放牧对环青海湖高寒草原主要植物种群生态位的影响.草业科学,2013,30(12),2040-2046.
Zheng W,Dong Q M,Li S X,Liu Y.Effects of grazing on niche of major plant populations in alpine steppe in Qinghai Lake Region.Pratacultural Science,2013,30(12):2040-2046.(in Chinese)
[23] 冯宜明,赵阳,陈学龙,齐瑞,秦燕燕,汤勇.甘肃亚高山云杉人工林抚育效果评价指标体系.兰州大学学报(自然科学版),2015,51(5):694-698.
Feng Y M,Zhao Y,Chen X L,Qi R,Qin Y Y,Tang Y.An evaluation index system for tending on spruce plantation at subalpine areas in Gansu Province.Journal of Lanzhou University(Natural Sciences),2015,51(5):694-698.(in Chinese)
[24] 赵栋,付作霖,徐德才,李丹春,齐瑞,齐昊.沙滩林场人工粗枝云杉林群落结构.水土保持研究,2017,24(1):346-350.
Zhao D,Fu Z L,Xu D C,Li D C,Qi R,Qi H.Community structure of planted picea asperata mast.forest in shatan forest farm.Research of Soil and Water Conservation,2017,24(1):346-350.(in Chinese)
[25] 周维.不同年龄云杉生物量和碳密度分布特征.长沙:中南林业科技大学硕士学位论文,2013.
Zhou W.Studies on biomass and carbon density in different-aged.Master Thesis.Changsha:Central South University of Forestry and Technology,2013.(in Chinese)
[26] 蔡慧颖,邸雪颖,金光泽.小兴安岭谷地云冷杉林的碳密度与生产力.应用生态学报,2014,25(10):2794-2802.
Cai H Y,Di X Y,Jin G Z.Carbon density and production in valley spruce-fir forest in Xiaoxing’an Mountains,China.Chinese Journal of Applied Ecology,2014,25(10):2794-2802.(in Chinese)
[27] 秦燕燕,蒋斌,曹秀文,冯宜明,李丹春,杨萌萌,向梅,陈蓉,李波.间伐强度对白龙江林区云杉人工林下植物多样性及其更新的影响.草业科学,2014,31(4):599-606.
Qin Y Y,Jiang B,Cao X W,Feng Y M,Li D C,Yang M M,Xiang M,Chen R,Li B.Effects of thinning intensity on diversity of undergrowth vegetation and regeneration in artifical spruce forest.Pratacultural Science,2014,31(4):599-606.(in Chinese)
[28] 冯宜明,车克钧,曹秀文,赵栋,谢飞,王杰.甘肃省白龙江林区主要森林类型生态服务功能价值评估.中南林业科技大学学报,2014,34(10):102-106.
Feng Y M,Che K J,Cao X W,Zhao D,Xie F,Wang J.Evaluation of eco-service functions of major forest types in Bailongjiang forest region of Gansu Province.Journal of Central South University of Forestry & Technology,2014,34(10):102-106.(in Chinese)
[29] 张金屯.数量生态学.北京:科学出版社,2004.
Zhang J T.Quantitative Ecology.Beijing:Science Press,2004.(in Chinese)
[30] 龚雪伟,吕光辉.艾比湖流域杜加依林荒漠植物群落多样性及优势种生态位.生物多样性,2017,25(1):34-45.
Gong X W,Lyu G H.Species diversity and dominant species’niches of eremophyte communities of the Tugai forest in the Ebinur basin of Xinjiang,China.Biodiversity Science,2017,25(1):34-45.(in Chinese)
[31] 戈峰.现代生态学.北京:科学出版社,2007.
Ge F.Modern Ecology.Beijing:Science Press,2007.(in Chinese)
[32] 高二鹏,杨海龙,贺康宁,曹恒,刘慧博,王冰洁.青海大通脑山区青海云杉+白桦混交林主要种群的生态位特征.中国水土保持科学,2014,12(3):101-106.
Gao E P,Yang H L,He K N,Can H,Liu H B,Wang B J.Niche characteristics of the main population underPiceacrassifoliaandBetulaplatyphyllamixed forest in high hill regions of Datong County,Qinghai Province.Science of Soil and Water Conservation,2014,12(3):101-106.(in Chinese)
[33] 王立龙,王广林,黄永杰,李晶,刘登义.黄山濒危植物小花木兰生态位与年龄结构研究.生态学报,2006,26(6):1862-1871.
Wang L L,Wang G L ,Huang Y J,Li J,Liu D Y.Age structure and niche of the endangeredMagnoliasieboldiiin Huangshan Mountain.Acta Ecologica Sinica,2006,26(6):1862-1871.(in Chinese)
[34] 张伟,何俊皓,郝文芳.黄土丘陵区不同管理方式下草地优势种群的生态位.草业科学,2016,33(7):1391-1402.
Zhang W,He J H,Hao W F.Niche characteristics of dominant plant populations in grassland of loess hilly region of China with different man-agement styles.Pratacultural Science,2016,33(7):1391-1402.(in Chinese)
[35] 卢炜丽,张洪江,陈奇伯,吴霖,栗宏林.重庆四面山杉木林群落乔木层优势种群生态位特征研究.西北林学院学报,2016,31(2):60-65.
Lu W L,Zhang H J,Chen Q B,Wu L,Li H L.Niche characteristics of the tree-layer dominant populations of cunninghamia lanceolata forest in chongqing simian mountains.Journal of Northwest Forestry University,2016,31(2):60-65.(in Chinese)
[36] 刘淑燕,余新晓,陈丽华.北京山区天然林乔木树种种间联结与生态位研究.西北林学院学报,2009,24(5):26-30.
Liu S Y,Yu X X,Chen L H.Interspecific association and niche research of natural forest in Beijing mountainous area.Journal of Northwest Forestry University,2009,24(5):26-30.(in Chinese)
[37] 周洋,郑小贤,王琦,刘晓玥.福建三明栲类次生林主要树种更新生态位研究.西北林学院学报,2015,30(4):84-88.
Zhou Y,Zheng X X,Wang Q,Liu X Y.Regeneration niche ofCastanopsissecondary forest in jiangle,Fujian.Journal of Northwest Forestry University,2015,30(4):84-88.(in Chinese)
[38] 刘巍,曹伟.长白山云冷杉群落主要种群生态位特征.生态学杂志,2011,30(8):1766-1774.
Li W,Cao W.Niche characteristics of main plant species in spruce-fir forests in Changbai Mountains.Chinese Journal of Ecology,2011,30(8):1766-1774.(in Chinese)
[39] 柴宗政,王得祥,张丽楠,张洋,黄青平,吴昊.秦岭山地天然油松群落主要植物种群生态位特征.生态学杂志,2012,31(8):1917-1923.
Chai Z Z,Wang D X,Zhang L N,Zhang Y,Huang Q P,Wu H.Niche characteristics of main plant populations in naturalPinustabulaeformis communities in Qinling Mountains,Northwest China.Chinese Journal of Ecology,2012,31(8):1917-1923.(in Chinese)
[40] 胡正华,钱海源,于明坚.古田山国家级自然保护区甜槠林优势种群生态位.生态学报,2009,29(7):3670-3677.
Hu Z H,Qian H Y,Yu M J.The niche of dominant species populations inCastanopsiseyreiforest in Gutian Mountain National Nature Reserve.Acta Ecologica Sinica,2009,29(7):3670-3677.(in Chinese)
[41] 陈艳瑞,尹林克.人工防风固沙林演替中群落组成和优势种群生态位变化特征.植物生态学报,2008,32(5):1126-1133.
Chen Y R,Yin L K.Community composition and niche change characteristics of dominant species in the wind-breaking and sand-fixing forest,Xinjiang,China.Journal of Plant Ecology,2008,32(5):1126-1133.(in Chinese)
