增温与氮沉降对高寒植物净光合速率的影响
2018-05-23张相锋董世魁赵金博赵珍珍韩雨晖杨明岳
冯 憬,张相锋,董世魁,赵金博,赵珍珍,韩雨晖,沙 威,杨明岳,李 帅,沈 豪
(北京师范大学环境学院,水环境模拟国家重点实验室,北京 100875)
21世纪全球平均气温将升高1.5~4.8 ℃[1],同时通过化石燃料燃烧和农业施肥沉降到陆地,生态系统的活性氮总量将增加两倍[2]。青藏高原正在经历显著的气候变暖[3],自1960年以来温度以每10年0.2 ℃增加[4]。预计在2030-2049年间,青藏高原气温将升高1.5 ℃,在2080-2099年气温将升高2.9 ℃[5]。研究表明,高纬度和高海拔地区(如青藏高原)的生态系统对气候变暖的响应可能更为敏感和迅速[6],气温升高不仅会改变高寒植物的光合和呼吸等生理过程[7],还导致群落种群高度、密度、盖度、频度、重要值[8]、土壤养分和水分利用效率的变化,如长期增温使高寒草甸植物群落高度增加、盖度降低[9-10],从而使植物的生长、生物量生产以及分配和群落演替发生了一定的变化[11]。另有研究表明,增温使物种丰富度减少,并使植物物种的多样性显著减少[12-13],同时会显著提高植被的地上净初级生产力[14]。
同时,青藏高原也面临氮沉降的严重影响。据报道,青藏高原氮沉降平均速率为8.7~13.8 kg·(ha·a)-1,已经显著改变了高原生态系统的生物化学循环[15]。氮作为植物的主要营养元素之一,氮沉降和外源氮素添加可以直接补充草地土壤速效养分,因而能够加快牧草的生长和发育,并改善草地群落的结构,提高牧草的产量[16-17]。青藏高原地区缺氮会造成物种大量丧失[12],人工施氮将在短期内影响高寒草地生态系统的物种组成、物种多样性以及生产力[18],可以使群落生物量尤其是优良牧草生物量增加[17,19]。前人对高原草地生态系统的研究工作表明,增温和氮沉降会改变物种种间关系从而影响植物群落的组成、结构和功能[20-22],其互作效应也将显著影响土壤碳、氮循环,如初级生产力[23-24]、微生物群落多样性[25]、凋落物分解及氮循环[26],进而影响到温室气体的排放,使之加剧。
迄今,前人的研究工作大多从中观或宏观尺度上揭示青藏高原草地生态系统对增温和氮沉降的响应,鲜有研究工作从植物生理生态等微观层面阐释增温和氮沉降对高寒草地植物的影响。因此,研究青藏高原增温及氮沉降背景下高寒草地植物的净光合速率变化,有助于揭示青藏高原高寒草地植物对全球变化的微观响应和反馈机制。基于这一背景,本研究以青藏高原地区的4类高寒草地——高寒草甸、高寒草原、高寒荒漠和栽培草地为研究对象,分别进行模拟增温、增氮处理以及增温和增氮耦合处理,以期证实科学假设:植物光合速率对增温和氮沉降的响应在草地类型和物种间存在差异,增温、氮沉降和二者的耦合效应对高寒草地植物光合速率的作用结果存在差异。
1 材料与方法
1.1 研究区概况
本研究在青藏高原典型区域青海省海北藏族自治州海晏县西海镇(36.93° N、100.95° E,海拔3 100 m)、青海省海南藏族自治中共和县铁卜加村(37.04° N、99.59° E,海拔3 270 m)以及新疆阿尔金山自然保护区即新疆巴音郭楞蒙古自治州若羌县祁曼塔格乡(37.30° N、90.34° E,海拔3 903 m)开展。西海镇年平均气温1.4 ℃,年均降水330~370 mm,草地植被类型以高寒草甸为主,优势种为线叶嵩草(Kobresiacapillifolia);铁卜加年均气温0 ℃,年均降水量377 mm,草地植被类型以高寒草原为主,优势种为紫花针茅(Stipacapillata);祁曼塔格乡年均气温1.64 ℃,年平均降水量17.4 mm,草地植被类型以高寒荒漠为主,优势种为猪毛菜(Salsolacollina)。
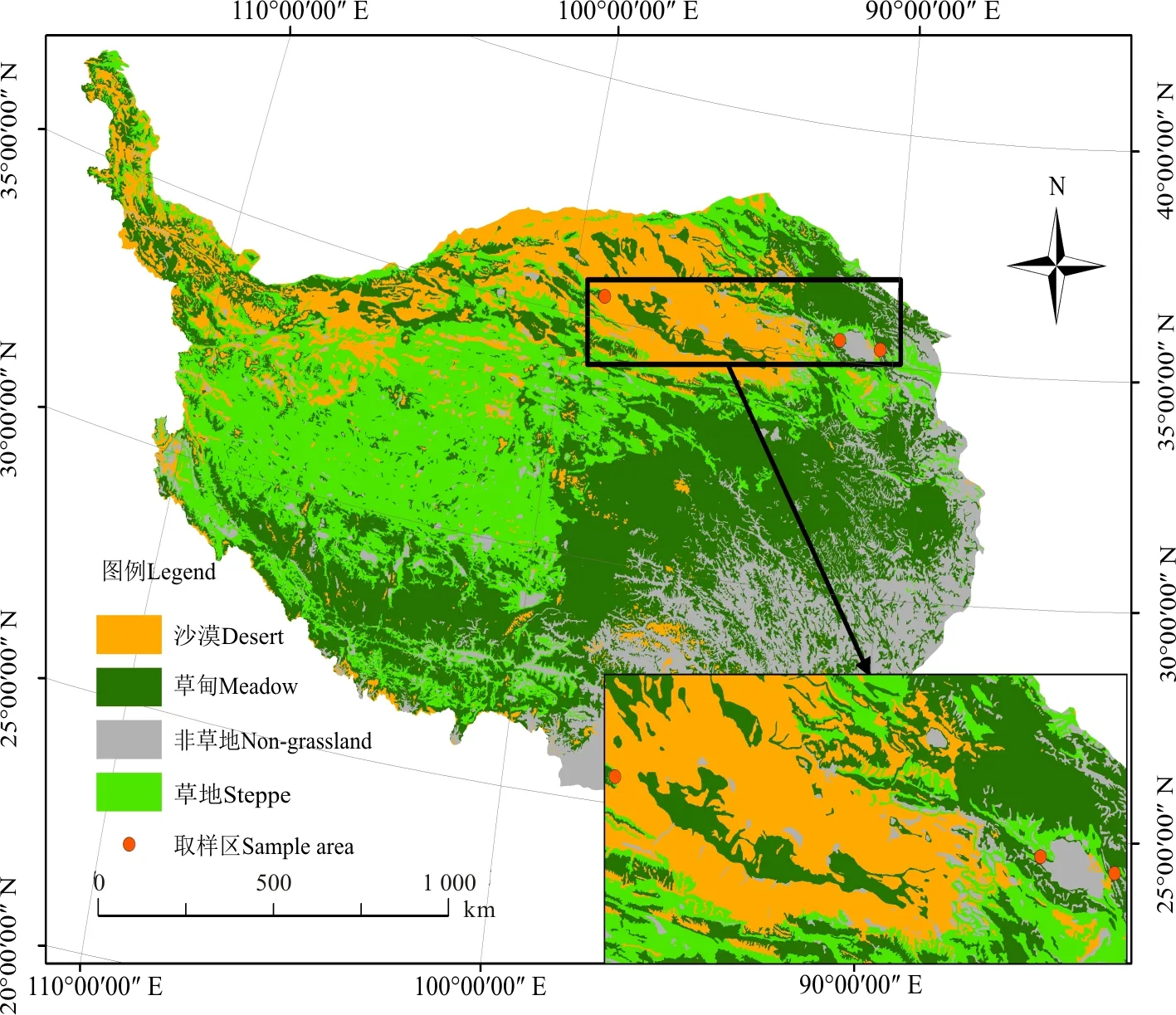
图1 研究区概况Fig. 1 Summary of the study area
1.2 样地设计
2014年9月在每个实验样地内布设6个圆台形开顶式增温室(truncated cone-shaped open top chamber ,OTC)(图2),使罩子内温度比外界温度高2 ℃以上,并用风扇调节实现罩子内均匀增温。2015年5月底随机选取3个OTC进行增温与施氮交互处理,氮肥采用硝酸铵(NH4NO3),施氮浓度为8 kg·(hm2·a)-1。同时,在3种草地上随机布设18个2 m×5 m的小区,随机选择3个为对照组(图3、图4),其余进行施氮处理,每个处理3次重复,施氮浓度分别为8、24、40、56、72 kg·(hm2·a)-1。海晏样地有7种不同的处理类型:CK、NⅠ、NⅡ、NⅢ、NⅣ、NⅤ、W、W-NⅠ,其中CK表示空白样地,NⅠ至NⅤ分别表示施氮处理浓度为8、24、40、56、72 kg·(hm2·a)-1,W表示增温样地,W-NⅠ表示增温和施氮耦合处理的样地,其中施氮浓度8 kg·(hm2·a)-1。铁卜加样地分为4种样地类型,即封育高寒草原、封育栽培草地(垂穗披碱草草地)、放牧高寒草原、放牧栽培草地(垂穗披碱草草地),其中封育高寒草原和封育栽培草地各有7种处理:CK、NⅠ、NⅡ、NⅢ、NⅣ、NⅤ、W、W-NⅠ,放牧高寒草原和放牧栽培草地均不做处理,以辅助解释围栏的作用。阿尔金山样地分为3种类型:盐渍化草原,荒漠与盐渍化草原过渡带,荒漠,由于试验条件限制该样地仅设增温处理:W、CK。
1.3 研究方法
在试验小区内选择生长状况良好的每种植物各3株,利用光合仪Li-6400测定植株第1或第2叶位叶片的净光合速率,叶室光强控制800 μmol·(m2·s)-1,测定时叶室为2 cm×3 cm,叶室光强为800 μmol·(m2·s)-1,叶室温度为25 ℃,叶室气体流速为500 μmol·(m2·s)-1,气孔比率为2,测得值为净光合速率。每种植物重复测3次,观测时间段选在7月中下旬,尽量选择天气晴好的日子进行,时间间隔为2~3 d,观测时间通常为09:00-11:00。本研究所采用的数据为2015年野外实地测量数据。对数据的异常值进行剔除,采用均值替换法,即用平均值来填充该缺失的变量值。
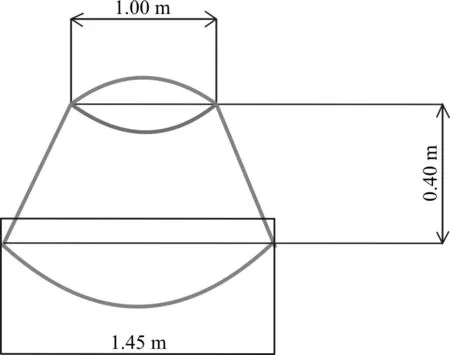
图2 圆台形开顶式增温室示意图(OTC)Fig. 2 Truncated cone-shaped open top chamber schematic diagram

图3 模拟增温、增温与氮沉降交互作用样地布局Fig. 3 Layout of sample areas on simulative warming and interactive treatment between warming and nitrogen adding
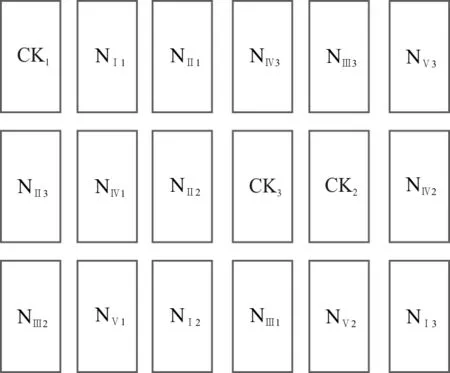
图4 模拟氮沉降试验小区布局Fig. 4 Layout of experimental sample areas on nitrogen adding
CK表示未处理样地即对照组,NⅠ至NⅤ分别表示施氮处理浓度为8、24、40、56、72 kg·(hm2·a)-1。
CK indicate control group, quadrat from NⅠto NⅤindicate adding nitrogen 8, 24, 40, 56, 72 kg·(hm2·a)-1, respectively.
1.4 数据分析
所有数据都在Excel 2016上处理后,用SPSS 16.0进行统计分析,采用单因素方差分析不同处理的效应,用Tukey检验对不同处理下植物净光合速率的差异性进行进一步检验(P<0.05),并用Origin 8.5作图。
2 结果与分析
2.1 高寒草甸优势植物净光合速率对增温和增氮的响应
在高寒草甸样地中,对照处理中白花枝子花(Dracocephalumheterophyllum)的平均净光合速率为2.24 μmol·(m2·s)-1(图5),仅在NⅡ处理下显著提高(P<0.05),而在其他增氮处理、增温处理及增温和增氮耦合处理下无显著变化(P>0.05),故白花枝子花属于低氮正向响应型植物,且对增温及增温和增氮耦合不产生响应;对照中的猪毛蒿(Artemisiascoparia)的平均净光合速率为0.15 μmol·(m2·s)-1,在NⅣ和NⅤ处理下显著升高,而对低氮、增温及增温和增氮耦合没有显著响应,属于高氮正向响应的植物;对照中的垂穗披碱草(Elymusnutans)、三出委陵菜(Potentillabetonicifolia)、唐松草(Thalictrumaquilegifolium)和狼毒(Euphorbiafischeriana)的平均净光合速率分别为2.70、1.59、1.61和0.26 μmol·(m2·s)-1,在不同梯度增氮、增温及增温和增氮耦合处理下均无显著性响应;冷地早熟禾(Poacrymophila)在对照处理下的平均净光合速率为1.04 μmol·(m2·s)-1,在全部增氮处理下均无显著变化,增温亦未显著改变其净光合速率,而增温和增氮耦合处理使其净光合速率显著升高;黄芪(Astragalusmembranaceus)在对照处理下的平均净光合速率为4.04 μmol·(m2·s)-1,在其他5组不同处理下其值均无显著变化,线叶嵩草(Kobresiacapillifolia)亦相同;针茅(Stipacapillata)在对照处理下的平均净光合速率为1.05 μmol·(m2·s)-1(图5),在NⅡ处理下其净光合速率显著升高;在对照样地中女娄菜(Sileneaprica)的净光合速率为1.34 μmol·(m2·s)-1,在NⅣ处理下显著增加,在其他4组增氮处理下无显著变化;矮生嵩草(Kobresiahumilis)在对照处理下的平均净光合速率为1.51 μmol·(m2·s)-1,在NⅣ和NⅤ处理下显著提高,在低氮和增温处理下均无显著变化;渐尖早熟禾(Poaattenuata)在对照处理下的平均净光合速率为0.95 μmol·(m2·s)-1,在NⅡ和NⅤ处理下显著提高,在NⅣ处理下显著骤增,在增温下无显著响应。
从高寒植物的光合作用响应特征来看,白花枝子花和针茅属于低氮正向响应型植物,即在低浓度增氮处理下净光合速率显著升高;猪毛蒿、女娄菜、矮生嵩草和渐尖早熟禾属于高氮正向响应的植物,即在高浓度增氮处理下净光合速率显著升高;冷地早熟禾属于对增温和增氮耦合处理正响应的植物类型;垂穗披碱草、三出委陵菜和唐松草对增氮均无显著性响应,此外,狼毒和线叶嵩草的净光合速率对低氮处理无显著响应,黄芪的净光合速率对高氮处理无显著响应,垂穗披碱草、黄芪和线叶嵩草对增温及增温和增氮耦合处理也无显著响应。

图5 海晏高寒草甸样地中植物净光合速率对增温和增氮的响应Fig. 5 Effect of simulative warming and nitrogen adding on net photosynthetic rate of alpine meadow plants on Haiyan
2.2 高寒草原优势植物净光合速率对增温和增氮的响应
在高寒草原样地中(图6),对照处理下的披针叶黄华(Thermopsislanceolata)的净光合速率为2.75 μmol·(m2·s)-1,在NⅢ处理下显著升高(P<0.05);马蔺在对照处理下的平均净光合速率为6.15 μmol·(m2·s)-1,仅在NⅡ和NⅤ处理下其净光合速率显著增加;矮生嵩草在对照处理下的平均净光合速率为1.55 μmol·(m2·s)-1,在NⅡ、NⅢ、NⅤ增氮处理及增温增氮耦合处理下其值无显著性变化(P>0.05);对照处理下,猪毛蒿的平均净光合速率为1.68 μmol·(m2·s)-1,在各组处理下均无显著性差异;多裂委陵菜(Potentillamultifida)在对照处理下的平均净光合速率为1.19 μmol·(m2·s)-1,在NⅤ增氮处理下有显著性提高;棘豆在对照处理下的平均净光合速率为1.03 μmol·(m2·s)-1,仅在NⅠ增氮处理下显著性增加,此外增温和增氮耦合对其也无显著性影响;线叶嵩草和冷地早熟禾在对照处理下的平均净光合速率分别为1.29和1.99 μmol·(m2·s)-1,在各处理下均无显著性差异;针茅在对照处理下的平均净光合速率为2.12 μmol·(m2·s)-1,在增温和不同浓度增氮处理下均无显著响应;甘肃马先蒿(Pediculariskansuensis)在对照处理下的平均净光合速率为2.99 μmol·(m2·s)-1,相较于对照组,其他各组处理均无显著差异,增温处理相比于NⅡ和NⅢ处理其净光合速率显著升高;平车前(Plantagodepressa)在对照处理下的平均净光合速率为3.28 μmol·(m2·s)-1,增氮处理对其并无显著影响;垂穗披碱草在对照处理下的平均净光合速率为6.15 μmol·(m2·s)-1,在NⅤ处理下显著升高,增温使其净光合速率显著降低,增温和增氮耦合对其无显著影响;冰草在对照处理下的平均净光合速率为5.50 μmol·(m2·s)-1,在NⅡ、NⅢ及增温和增氮耦合处理下显著降低;白花枝子花在对照处理下的平均净光合速率为2.57 μmol·(m2·s)-1,在低氮处理及增温增氮耦合处理下其值无显著性变化。
在栽培草地中(图7),披针叶黄华、垂穗披碱草、多枝黄芪、冷地早熟禾、猪毛蒿和紫菀(Astragalusmembranaceus)的平均净光合速率分别为 5.08、5.11、4.94、1.83、2.02和3.48 μmol·(m2·s)-1。披针叶黄华属于对增氮负向响应的植物,在4个增氮处理中,其净光合作用均低于对照组;垂穗披碱草属于低氮浓度响应型植物,在NⅠ增氮样地中其净光合速率显著升高(P<0.05),在其他增氮处理中均显著降低;多枝黄芪属于对增氮负向响应的植物类型,5组增氮处理均使其净光合速率降低,其中NⅢ和NⅣ处理使其显著降低;冷地早熟禾(Poacrymophila)属于低氮浓度响应型植物,在1倍增氮样地中其净光合速率显著升高,在NⅡ和NⅢ处理下显著降低;猪毛蒿属于高氮响应型植物,仅在NⅤ处理下其净光合速率显著升高;紫菀属于对增氮负向响应的植物,在NⅡ处理下其净光合速率显著降低,在其他处理下无显著性差异(P>0.05)。
2.3 高寒荒漠优势植物净光合速率对增温的响应
在盐渍化草原中(图8),围栏内对照样地的西伯利亚蓼(Polygonumsibiricum)、红景天(Rhodiolarosea)、华扁穗草(Blysmussinocompressus)和赖草(Leymussecalinus)的平均净光合速率分别为2.75、0.81、2.66和3.04 μmol·(m2·s)-1。相较于对照组,增温使华扁穗草的净光合速率显著降低(P<0.05),赖草的净光合速率降低,但不显著,红景天和西伯利亚蓼的净光合速率无显著变化(P>0.05)。
在高寒荒漠中,围栏内对照样地的垫状驼绒藜(Ceratoidescompacta)、黄花补血草(Limoniumaureum)、针茅和蒿叶猪毛菜(Salsolaabrotanoides)的净光合速率分别为1.54、1.40、0.60和1.68 μmol·(m2·s)-1(图8)。相较于对照组,增温使垫状驼绒藜的净光合速率显著增加(P<0.05),黄花补血草和蒿叶猪毛菜的净光合速率降低,但不显著(P>0.05),针茅无显著变化。
在高寒荒漠与盐渍化草原的过渡带中,围栏内对照样地的赖草、粗壮嵩草(Kobresiarobusta)、蒿叶猪毛菜和针茅的平均净光合速率分别为1.16、2.55、2.09和0.827 μmol·(m2·s)-1(图8)。相较于对照组,模拟增温使赖草的净光合速率显著增加(P<0.05),而粗壮嵩草、针茅和蒿叶猪毛菜的净光合速率均无显著变化(P>0.05)。
3 讨论与结论
研究结果验证了科学假设,即植物光合速率对增温和氮沉降的响应在草地类型和物种间存在差异,增温、氮沉降和二者的耦合效应对高寒草地植物的光合速率的作用结果存在差异。有研究表明,缺氮能够显著降低植物叶绿素含量和光合速率,而施加一定浓度的氮能够增加植物的净光合速率[27]。但同时植物对氮浓度也有一定的耐受范围,对植物来说当氮沉降量超出耐受范围时必将影响其生长,最直观可以体现在净光合速率等光合指标上[28]。增温对植物的影响也类似,但是不同植物对增温和增氮的响应是不同的。开顶式增温罩通过增温改善了植物群落生长局部环境,在一定程度上促进了植物的生长,短时内可以促进绝大数禾本科植物生长,由于其对于光照利用的竞争较强,所以相对高度和盖度都将增加,同时相关研究还表明,温度升高植物的生长期会延长,这有利于草地植物生物量的增加[29]。此外,增温和增氮耦合处理对大部分植物净光合速率的提升有显著作用。
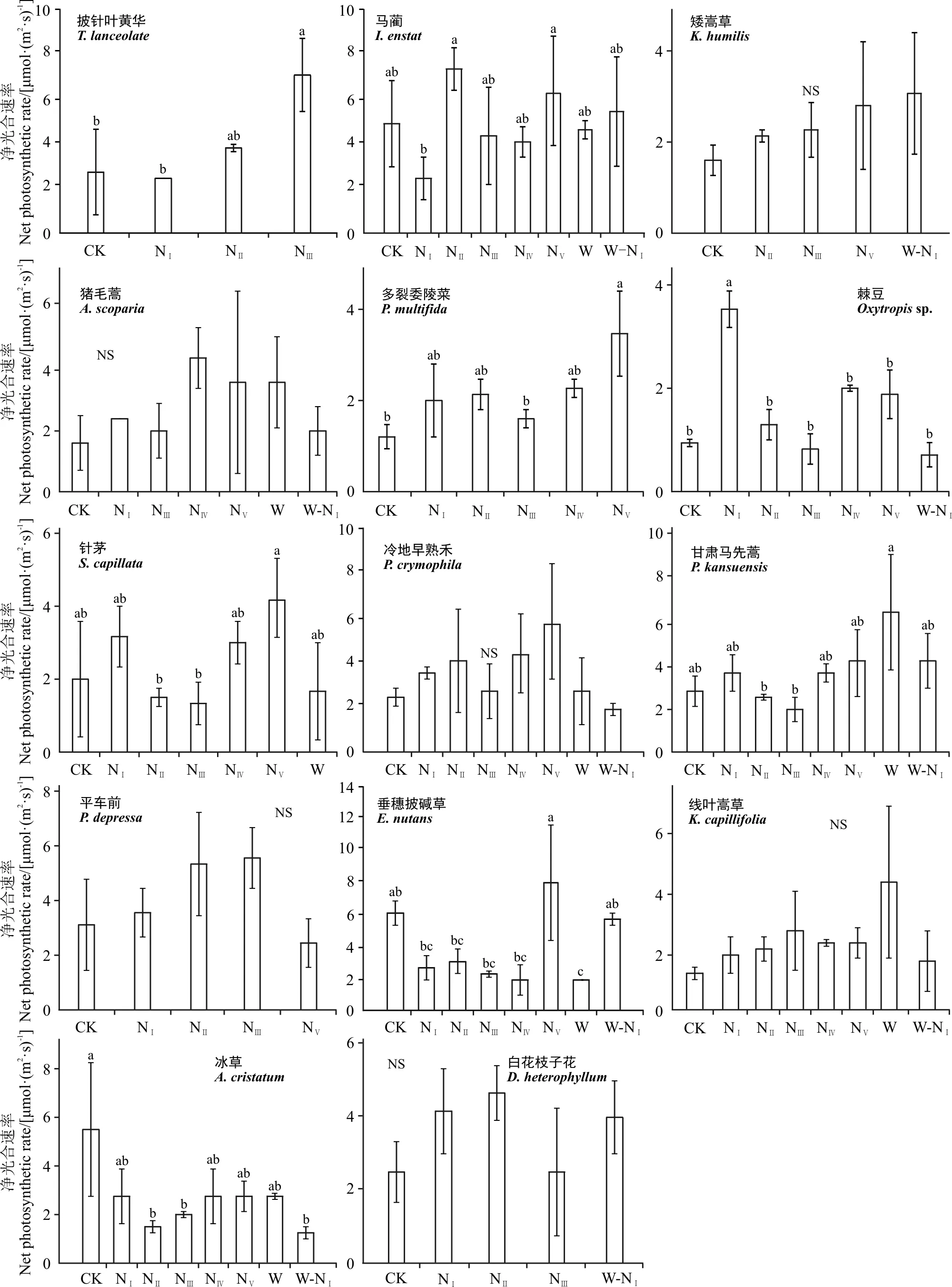
图6 铁卜加自然草地植物净光合速率对增温和增氮处理的响应Fig. 6 Effect of simulative warming and nitrogen adding on net photosynthetic rate of alpine natural grassland plants on Tiebujia

图7 铁卜加栽培草地植物净光合速率对增氮处理的响应Fig. 7 Effect of simulative warming and nitrogen adding on net photosynthetic rate of alpine artificial grassland plants on Tiebujia

图8 阿尔金山保护区盐渍化草原、荒漠以及荒漠和盐化草原过渡带3种样地中植物净光合速率对增温的响应Fig. 8 Effect of simulative warming and control treatment on net photosynthetic rate of salinization grassland, alpine desert and transitional zone plants on Aerjin mountain natural reserve areas
不同小写字母表示增温与对照间差异显著(P<0.05)。
Different lowercase letters indicate significant differences between warming and control at the 0.05 level.
通过本研究,可以从植物光合作用这一微观尺度得出如下结论,在高寒草甸中,白花枝子花和针茅属于低氮正向响应型植物,即在低浓度增氮处理下净光合速率显著升高;猪毛蒿、女娄菜、矮生嵩草和渐尖早熟禾为高氮正向响应的植物,即在高浓度增氮处理下净光合速率显著升高;冷地早熟禾为对增温和增氮耦合处理正响应的植物类型;在高寒草原中,棘豆为低氮正向响应型植物,披针叶黄华为中氮正向响应型植物,多裂委陵菜马蔺、猪毛蒿、冷地早熟禾、甘肃马先蒿、线叶嵩草为对增氮不显著的植物,针茅为对增温和增氮均不响应的植物,平车前为对增氮不响应的植物,垂穗披碱草为增温负向响应的植物,冰草为低氮中氮负向响应型植物以及增温和增氮耦合负向响应型植物;在栽培草地中,披针叶黄华为高氮负向响应型植物,垂穗披碱草为低氮正向响应型植物兼中氮负向响应型植物,多枝黄芪为中氮高氮负向响应型植物,冷地早熟禾为中氮负向响应型植物,猪毛蒿为高氮正向响应型植物,紫菀为低氮负向响应型植物;在高寒荒漠中,华扁穗草为增温负向响应型植物,垫状驼绒藜为增温正向响应型植物,而红景天、西伯利亚蓼、赖草、黄花补血草、针茅、蒿叶猪毛菜以及粗壮嵩草均为增温不显著响应类型的植物。
本研究仅采用了净光合速率作为分析指标,在今后对于高寒植物生理生态对增温和氮沉降的响应规律研究中,应增加更多与植物光合相关的指标进行全面分析,如蒸腾速率、气孔导度和水分利用率等,同时高寒荒漠样地也应增设不同梯度施氮试验,以全面揭示高寒植物在生理生态微观尺度上对增温和氮沉降的响应。
参考文献References:
[1] Stocker T,Qin D,Plattner G,Tignor M,Allen S,Boschung J,Nauels A,Xia Y,Bex V,Midgley P.IPCC 2013:Climate Change 2013:The Physical Science Basis.Contribution of Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change.Cambridge,United Kingdom and New York:Cambridge University Press,2013:1535.
[2] Galloway J N,Dentener F J,Capone D G,Boyer E W,Howarth R W,Seitzinger S P,Asner G P,Cleveland C C,Green P A,Holland E A,Karl D M,Michaels A F,Porter J H,Townsend A R,Vörösmarty C J.Nitrogen Cycles:Past,present,and future.Biogeochemistry,2004,70(2):153-226.
[3] Gao Q Z,Ganjurjav,Li Y,Wan Y F,Zhang W N,Almaz Borjigdai.Challenges in disentangling the influence of climatic and socio-economic factors on alpine grassland ecosystems in the source area of Asian major rivers.Quaternary International,2013,304(9):126-132.
[4] Chen H,Zhu Q,Peng C,Wu N,Wang Y F,Fang X Q,Gao Y H,Zhu D,Yang G,Tian J Q,Kang X M,Piao S L,Ouyang H,Xiang W H,Luo Z B,Jiang H,Song X Z,Zhang Y,Yu G R,Zhao X Q,Gong P,Yao T D,Wu J H.The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau.Global Change Biology,2013,19(10):2940-2955.
[5] Liu X D,Cheng Z G,Yan L B,Yin Z Y.Elevation dependency of recent and future minimum surface air temperature trends in the Tibetan plateau and its surroundings.Global & Planetary Change,2009,68(3):164-174.
[6] 布仁巴音,徐广平,段吉闯,常小峰,张振华,汪诗平.青藏高原高寒草甸初级生产力及其主要影响因素.广西植物,2010(6):760-769.
Burenbayin,Xu G P,Duan J C,Chang X F,Zhang Z H,Wang S P.Primary productivity and its main affecting factors of alpine meadows on the Tibetan plateau.Guihaia,2010(6):760-769.(in Chinese)
[7] 珊丹,韩国栋,赵萌莉,王珍,韩雄,高福光.控制性增温和施氮对荒漠草原土壤呼吸的影响.干旱区资源与环境,2009,23(9):106-112.
Shan D,Han G D,Zhao M L,Wang Z,Han X,Gao F G.The effects of experimental warming and nitrogen addition on respiration in desert steppe.Journal of Arid and Resources and Environment,2009,23(9):106-112.(in Chinese)
[8] 李娜,王根绪,杨燕,高永恒,柳林安,刘光生.短期增温对青藏高原高寒草甸植物群落结构和生物量的影响.生态学报,2011,31(4):895-905.
Li N,Wang G X,Yang Y,Gao Y H,Liu L N,Liu G S.Short-term effects of temperature enhancement on community structure and biomass of alpine meadow in the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2011,31(4):895-905.(in Chinese)
[9] Zhang Y,Gao Q Z,Dong S K,Liu S L,Wang X X,Su S K,Li Y Y,Tang L,Wu X Y,Zhao H D.Effects of grazing and climate warming on plant diversity,productivity and living state in the alpine rangelands and cultivated grasslands of the Qinghai-Tibetan Plateau.Rangeland Journal,2015,37(1):57-65.
[10] 赵艳艳,周华坤,姚步青,王文颖,董世魁,赵新全.长期增温对高寒草甸植物群落和土壤养分的影响.草地学报,2015,23(4):665-671.
Zhao Y Y,Zhou H K,Yao B Q,Wang W Y,Dong S K,Zhao X Q.The influence of long-term simulating warming to the plant community and soil nutrient of alpine meadow.Acta Agrestia Sinica,2015,23(4):665-671.(in Chinese)
[11] Henry G H R,Molau U.Tundra plants and climate change:The International Tundra Experiment (ITEX).Global Change Biology,1997,3(S1):1-9.
[12] Klein J A,Harte J,Zhao X Q.Experimental warming causes large and rapid species loss,dampened by simulated grazing,on the Tibetan Plateau.Ecology Letters,2004,7(12):1170-1179.
[13] Klein J A.Climate change and its ecological impacts:The evidences from the field experiments on northeastern Tibetan plateau.//Abstracts of EcoSummit 2007 Ecological Complexity and Sustainability:Challenges & Opportunities for 21st Century’s Ecology,2007.
[14] Wang S P,Duan J C,Xu G P,Wang Y F,Zhang Z H,Rui Y C,Luo C Y,Xu B R B Y,Zhu X X,Chang X F,Gui X Y,Niu H S,Zhao X Q,Wang W Y.Effects of warming and grazing on soil N availability,species composition,and ANPP in an alpine meadow.Ecology,2012,93(11):2365-2376.
[15] Lyu C,Tian H.Spatial and temporal patterns of nitrogen deposition in China:Synthesis of observational data.Journal of Geophysical Research-Atmospheres,2007,112(D22):229-238.
[16] 郭江松,于磊,鲁为华,林祥群.施肥对驯化条件下线叶野豌豆饲草和种子生产的影响.草原与草坪,2010,30(1):61-63.
Guo J S,Yu L,Lu W H,Lin X Q.Effect of fertilization on forage yield and seed production ofViciatenuifolia.Grassland and Turf,2010,30(1):61-63.(in Chinese)
[17] 郭红玉.模拟增温和氮素添加对高寒草甸草地的影响.西宁:青海大学硕士学位论文,2015.
Guo H Y.The impact of the simulative warming and adding nitrogen on grassland in alpine meadow.Master Thesis.Xining:Qinghai University,2015.(in Chinese)
[18] 杨元武,周华坤,李希来,周旭辉,高健军,刘玉,赵新全,叶鑫.高寒草甸物种多样性和生产力对养分添加的初期响应.西北农业学报,2017,26(2):159-166.
Yang Y W,Zhou H K,Li X L,Zhou X H,Gao J J,Liu Y,Zhao X Q,Ye X.Initial response of species diversity and productivity to nutrients addition on alpine meadow.Acta Agriculturae Boreali-occidentalis Sinica,2017,26(2):159-166.(in Chinese)
[19] 郭雅婧.氮素添加对青藏高原高寒草甸植被和土壤氮素供应能力的影响.兰州:兰州大学硕士学位论文,2015.
Guo Y J.Effects of nitrogen adding on plant community and soil nitrogen supply ability of alpine meadow in Qinghai-Tibet Plateau.Master Thesis.Lanzhou:Lanzhou University,2015.(in Chinese)
[20] Harte J,Shaw R.Shifting dominance within a montane vegetation community:Results of a climate-warming experiment.Science,1995,267:876880.
[21] Yang H J,Wu M Y,Liu W X,Zhang Z,Zhang N L,Wan S Q.Community structure and composition in response to climate change in a temperate steppe.Global Change Biology,2015,17(1):452-465.
[22] 李元恒.内蒙古荒漠草原植物群落结构和功能对增温和氮素添加的响应.呼和浩特:内蒙古农业大学博士学位论文,2014.
Li Y H.Responses of plant community structure and function to warming and nitrogen addition in a desert steppe of Inner Mongolia.PhD Thesis.Huhhot:Inner Mongolia University,2014.(in Chinese)
[23] Melillo J M,Cowling E B,刘林群.活性氮与环保公共政策.AMBIO-人类环境杂志,2002,31(2):150-158.
Melillo J M,Cowling E B,Liu L Q.Reactive nitrogen and public policies for environmental protection.AMBIO-A Journal of the Human Environment,2002,31(2):150-158.(in Chinese)
[24] Shaver G R,Chapin F S,Gurevitch J,Harte J,Henry G Ineson P,Jonasson S,Melillo J,Pitelka L,Rustad L,Canadell J.Global warming and terrestrial ecosystems:A conceptual framework for analysis.BioScience,2000,50(10):871-882.
[25] Frey S D,Drijber R,Smith H,Melillo J.Microbial biomass,functional capacity,and community structure after 12 years of soil warming.Soil Biology & Biochemistry,2008,40(11):2904-2907.
[26] Berg B,Mcclaugherty C,Johansson M B.Litter mass-loss rates in late stages of decomposition at some climatically and nutritionally different pine sites.Long-term decomposition in a Scots pine forest.VIII.Canadian Journal of Botany,1993,71(5):680-692.
[27] 郭二辉,胡聃,田朝阳,胡颖,王从彦,于盈盈.土壤氮素与水分对植物光合生理生态的影响研究.安徽农业科学,2008,36(26):11211-11213.
Guo E H,Hu D,Tian C Y,Hu Y,Wang C Y,Yu Y Y.Study on the effects of soil nitrogen and moisture on plant photosynthetic physiological ecology.Journal of Anhui Agricultural Sciences,2008,36(26):11211-11213.(in Chinese)
[28] 周晓兵,张元明,王莎莎,张丙昌,张静.3种荒漠植物幼苗生长和光合生理对氮增加的响应.中国沙漠,2011,31(1):82-89.
Zhou X B,Zhang Y M,Wang S S,Zhang B C,Zhang J.Effect of nitrogen input on growth and photosynthetic physiology of three desert species seedlings.Journal of Desert Research,2011,31(1):82-89.(in Chinese)
[29] 周华坤,周兴民,赵新全.模拟增温效应对矮生嵩草草甸影响的初步研究.植物生态学报,2000,24(5):547-553.
Zhou H K,Zhou X M,Zhao X Q.A preliminary study of the influence of simulated greenhouse effect on aKobresiahumilismeadow.Acta Phytoecologica Sinica,2000,24(5):547-553.(in Chinese)
