金银花害虫咖啡脊虎天牛对三种寄主的产卵行为研究△
2018-05-21陈建民刘赛乔海莉徐常青徐荣郭昆陈君
陈建民,刘赛,乔海莉,徐常青,徐荣,郭昆,陈君
(中国医学科学院 北京协和医学院 药用植物研究所,北京 100193)
金银花Lonicerajaponica为忍冬科植物忍冬的干燥花蕾或初开花。性甘、寒。归肺、心、胃经。具有清热解毒、疏散风热的功效。用于治疗痈肿疔疮,喉痹,丹毒,热毒血痢,风热感冒,温病发热等症状[1]。广泛应用于药品、食品、化妆品和保健品等行业[2]。咖啡脊虎天牛Xylotrechusgrayii属鞘翅目天牛科天牛亚科,是我国金银花产区的重要害虫。主要分布于中国、日本、韩国,在中国广泛分布于华北、甘肃、西藏、四川、江苏、福建、广东、台湾。报道的寄主植物有金银花Lonicerajaponica、咖啡Coffeaarabica、柚木Tectonagrandis、榆Ulmus、泡桐Pauloniatomentosa、山石榴Randiaspinosa、漆树Odina、刺楸Kalopanax、赤楠Syzygiumbuxifolium、冷杉Abies、柑橘Citrus、香榧Torreyagrandis等[3-6]。目前有关咖啡脊虎天牛危害寄主以金银花报道最多,其它寄主少有报道。咖啡脊虎天牛以幼虫蛀食金银花枝干韧皮部和木质部,导致金银花枝干枯萎甚至整株死亡,发生严重地区金银花死亡率高达70-80%,目前,此害虫有扩散加重趋势[7-11]。笔者调查显示,在山东平邑金银花产区主要有3种寄主植物,分别是金银花、泡桐和榆树,而咖啡脊虎天牛仅危害金银花,周边的泡桐和榆树未见危害。本文以金银花、泡桐和榆树为试验材料,通过行为学观察等方法,比较不同发育阶段咖啡脊虎天牛对3种寄主的选择差异,以期探索其寄主选择机制,为咖啡脊虎天牛防治提供理论依据。
1 材料和方法
1.1 供试材料
咖啡脊虎天牛(成虫和幼虫):分别于2017年2月至7月采集自山东省临沂市平邑县金银花产区,带回北京室内8℃冷藏备用。
未交尾成虫,将野外采集的未交尾成虫自冷藏室中取出进行性别鉴定,然后分别放置于加入湿棉球的培养皿中,在室温下恢复24 h后备用。
交尾成虫,将野外采集的未交尾成虫在室温下交尾,选择首次交尾后一天的健壮雌雄成虫进行测定。
金银花、泡桐和榆树:采集自药用植物研究所药用植物园。
1.2 试验方法
1.2.1 成虫交尾前后对不同寄主的选择 剪取30 cm长的新鲜金银花、泡桐和榆树枝条,分别插入装水的锥形瓶中,并用锡箔纸将枝条和瓶口包裹固定。设置一个1 m×1 m×1.5 m的纱网观察笼,在底部选择一个最大的等边三角形区域,将以上3种枝条分别放置在这个区域的3个角。将待测成虫自等边三角形区域的中心释放,每次释放1只成虫,观察30 min,首次到达某一枝条记为选择。每测试3只,顺时针变换一次枝条位置。每个处理至少测定9只,重复3次。前期研究发现成虫白天活动,且在9∶00~17∶00活动旺盛,故测试选择在这一时段。
1.2.2 交尾雌性成虫对不同寄主的产卵选择 剪取长度10 cm、直径相同的新鲜泡桐、榆树、金银花枝条各3段,放入直径20 cm的培养皿中,重复10组。将培养皿放入光周期16∶8、温度28 ℃∶25 ℃、湿度40%的培养箱(宁波江南仪器厂RXZ-380A型培养箱)。每个培养皿中释放一对交尾后的雌雄成虫,释放3天后开始观察,每天观察一次,观察至雌性成虫停止产卵,记录产卵量、产卵位置及位置结构。每天更换一次枝条,并将原枝段分树种放入新的培养皿,在同样条件下观察卵的孵化和一龄幼虫的取食。
1.2.3 幼虫在不同寄主上的取食和羽化 幼虫的取食和羽化实验在培养箱中进行,条件同上。选取同一时间野外采集的大小相同健康状况良好的幼虫20只,采集直径相同的金银花和泡桐枝条,将其剪成6 cm长的枝段,每一树种选取10段,分别放入10 cm的玻璃指形管,并在其中放入一只上述幼虫,每个指形管用脱脂棉做塞,塞棉上滴适量水,以保证管内湿度,重复3次。每3天检查一次,每次检查时更换干枯枝段,前4次观察时,记录每个树种幼虫取食数量,并称量每只幼虫产生的虫屑量。观察至幼虫死亡或蛹羽化,统计羽化率。
1.2.4 交尾雌性成虫在不同金银花枝段的产卵选择 实验在培养箱内进行,条件设置同上。将首次交尾的雌雄成虫和不同处理的金银花枝段放入直径10 cm的培养皿,每个培养皿放置1对成虫和不同处理金银花枝段各3段,重复10次。每天观察,记录产卵量、产卵位置,并更换相应枝段,连续观察至雌虫停止产卵。不同处理金银花枝段如下:光滑金银花枝段,选择表面光滑的金银枝条,将其切成等长的枝段,将两端用石蜡封口;带缝隙金银花枝段,用解剖刀在光滑金银花枝段表面纵向划出三条长20 mm宽1~2 mm的缝隙。带切面金银花枝段,用解剖刀将光滑金银花枝段表面切出长20 mm宽2~3 mm的光滑切面。
1.3 分析方法
运用SPSS 20.0( IBM,Chicago,IL)软件进行数据分析。对不同寄主的定向和产卵数据选择单因素方差分析,对纱网和寄主的选择数据及卵的孵化率使用T检验分析。
2 结果与分析
2.1 成虫交尾前后对不同寄主的选择
交尾前后的两性成虫对寄主和纱网有显著的选择差异(交尾雄t=3.881,df=4,P<0.05,n=30;交尾雌t=3.618,df=4,P<0.05,n=28;未交尾雄t=6.124,df=4,P<0.05,n=27;未交尾雌t=16.971,df=4,P<0.05,n=28),大量成虫选择在纱网上停留。成虫对不同寄主的选择率如图1。未交尾的雄性成虫不选择榆树,对金银花和泡桐的选择率分别为3.7%和18.5%,77.8%选择观察笼的纱网。未交尾的雌性成虫也不选择榆树,对金银花和泡桐的选择率分别为3.7%和3.3%,93.0%选择观察笼纱网。交尾后的雄性成虫不选择金银花和榆树,对泡桐和观察笼纱网的选择率分别为22.0%和78.0%。交尾后的雌性成虫对金银花、泡桐和榆树的选择率分别为3.3%、18.2%和7.4%,仍有71.1%选择观察笼的纱网。交尾前后的两性成虫对金银花、泡桐和榆树均没有显著的选择差异(未交尾雄F=4.200,df=2,P>0.05,n=27;未交尾雌F=0.500,df=2,P>0.05,n=28;交尾雄F=3.769,df=2,P>0.05,n=30;交尾雌F=2.167,df=2,P>0.05,n=28)。
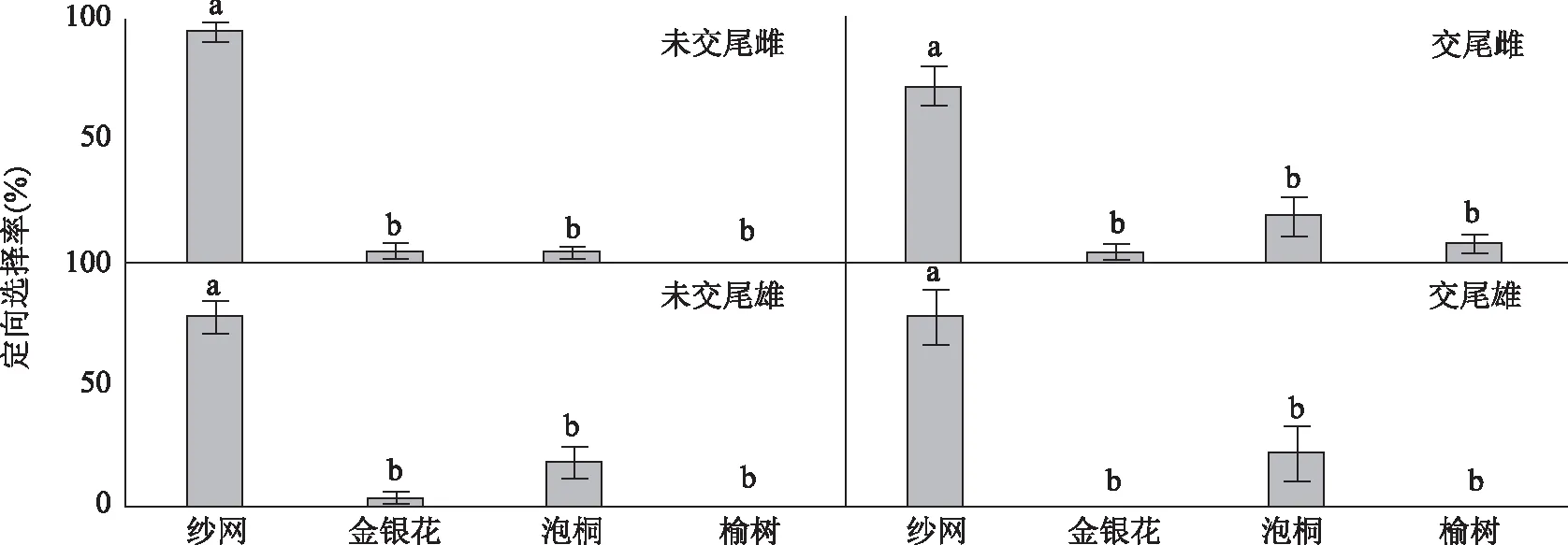
注:不同字母表示单因素方差分析在0.05水平上差异显著。图1 咖啡脊虎天牛成虫对不同寄主的定向选择率
2.2 交尾雌性成虫对不同寄主的产卵选择
交尾雌虫在榆树枝条上未见产卵,在金银花和泡桐上的平均产卵量和孵化率如表1,在金银花上的产卵量显著多于泡桐(t=2.710,df=9,P<0.05,n=10)。雌虫大量卵产于金银花切口缝隙处、表皮裂缝或皮下缝隙,少量产于节间缝隙,少有几粒卵产于金银花枝段表面,未发现刻槽产卵现象。在金银花和泡桐上卵的孵化率无显著差异(t=0.360,df=3.442,P>0.05,n=10)。形态正常的卵,在金银花和泡桐上均能孵化,且一龄幼虫能够取食嫩皮后向内蛀入。
表1 咖啡脊虎天牛在不同寄主上的产卵量和孵化率

表1 咖啡脊虎天牛在不同寄主上的产卵量和孵化率
注:不同字母表示同列不同行间在0.05水平上差异显著。
2.3 幼虫在不同寄主上的取食和羽化
幼虫取食枝段横截面或者钻入枝段与管壁形成的缝隙从枝段表面向内蛀食。幼虫对金银花和泡桐的平均选择率分别为53.3%和69.2%(表2)。幼虫在金银花和泡桐上取食3天所产生的平均虫屑量分别为40.9 mg和59.2 mg(表2),在两种寄主之间幼虫选择率和虫屑量均无显著差异(选择率t=0.700,df=22,P>0.05,n=12;虫屑量t=1.214,df=137,P>0.05,n=120)。取食两种寄主的幼虫均能羽化,羽化率无显著差异(t=2.012,df=4,P>0.05,n=3)。
表2 咖啡脊虎天牛幼虫在不同寄主上的取食选择率、虫屑量和羽化率
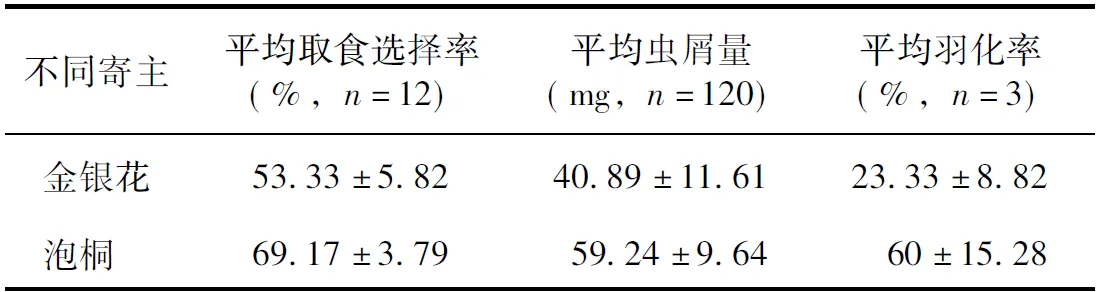
表2 咖啡脊虎天牛幼虫在不同寄主上的取食选择率、虫屑量和羽化率
2.4 交尾雌性成虫在不同金银花枝段的产卵选择
雌性成虫在表面光滑金银花枝段上的平均产卵量是0.5粒,在带切面金银花枝段的平均产卵量是5.5粒,在带缝隙金银花枝段的平均产卵量是24.2粒。雌性成虫在带缝隙枝段的产卵量显著多于在光滑枝段和带切面枝段的产卵量(缝隙与光滑t=11.282,df=9,P<0.05,n=10;缝隙与切面t=5.442,df=9,P<0.05,n=10),而产卵量在光滑枝段和带切面枝段之间无显著差异(t=2.177,df=9,P>0.05,n=10)(图2)。
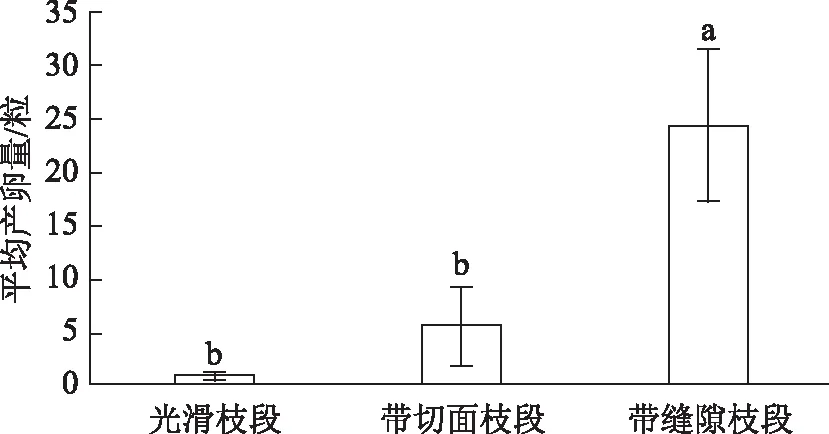
注:不同字母表示单因素方差分析在0.05水平上差异显著。图2 雌性成虫在不同表面结构金银花枝段上的平均产卵量
3 讨论
天牛在寄主选择过程中分为多个阶段,从成虫的定向、产卵到卵的孵化、幼虫的取食和成虫羽化,每个阶段都伴随天牛对寄主的选择。咖啡脊虎天牛成虫对寄主无明显的选择趋向,且不能区分金银花、泡桐和榆树。这与其它天牛有很大差异。野蔷薇和杨树能够显著的吸引杨树云斑天牛,松墨天牛Monochamusalternatus对寄主有明显的趋性,国槐枝条和叶片对锈色粒肩天牛Aprionaswainsoni的吸引力明显大于香花槐、栾树、黄山栾、黄檀和云实的枝条和叶片[12-14]。诸多研究发现,许多天牛主要依靠嗅觉和视觉来定位寄主,且能够根据寄主的颜色和气味识别寄主[15-20],本研究结果显示咖啡脊虎天牛进行寄主选择和定位可能并非主要依靠嗅觉和视觉。
然而,咖啡脊虎天牛在产卵阶段的寄主选择中,雌性成虫能够明显区分金银花、泡桐和榆树,显著的选择金银花,大量卵产于金银花枝条缝隙内,无刻槽产卵行为。研究发现,榆树和泡桐枝条表面光滑,很少有老皮翘起,无明显缝隙结构,这可能是雌性成虫无法产卵的重要原因。而金银花的生长会造成老皮开裂并逐渐与内层分离形成间隙,这为雌性成虫的产卵提供了有效的缝隙结构。雌性成虫对不同金银花枝段的产卵结果,进一步证实缝隙结构是其确定产卵位置的重要因素。以往研究发现,许多有刻槽产卵的天牛都有补充营养的行为,如梣天牛Eutetraphasedecimpunctata、桑天牛Aprionagermari、云斑白条天牛Batoceralineolata、橙斑白条天牛Batoceradavidia、双斑锦天牛Acaloleptasublusca、梨眼天牛BacchisaFortunei、榕八星天牛Batocerarubus等[21-27]。而不进行刻槽产卵的天牛多不补充营养或仅取食少量水液,如栎旋木柄天牛Aphrodisiumsauteri、园斑紫天牛Purpuricenussideriger、花椒虎天牛Clytusvalidus、桑虎天牛Xylotrechuschinensis等[28-31]。这与咖啡脊虎天牛的行为一致。可能也是导致其成虫对寄主没有明显选择趋性的重要原因。
本研究发现,咖啡脊虎天牛成虫对于寄主无明显的定向行为,寄主的颜色和气味不会引起成虫对寄主的定向选择差异。咖啡脊虎天牛雌性成虫通过触觉选择确定产卵场所是造成其寄主选择差异的主要原因,寄主表面的缝隙结构是产卵选择的重要特征。在卵的孵化、幼虫取食和羽化阶段,饱满的卵在金银花和泡桐上均能孵化,无显著差异;幼虫均能取食金银花和泡桐,羽化率亦无显著差异。这些结果显示控制育苗过程中的卵和幼虫携带、去除寄主枝干表面的产卵结构、扰乱成虫的交尾行为都可以有效的控制咖啡脊虎天牛的扩散和危害。
[1] 国家药典委员会.中华人民共和国药典:一部[S].北京:中国医药科技出版社,2015:221.
[2] 王亚丹,杨建波,戴忠,等.中药金银花的研究进展[J].药物分析杂志,2014,34(11):1928-1935.
[3] 陈世骧,谢蕴贞,邓国藩.中国经济昆虫志第一册鞘翅目天牛科[M].北京:科学出版社.1959:57-58.
[4] 蒋书楠.中国天牛幼虫[M].重庆:重庆出版社.1989:116.
[5] Han Y,Lyu D.Taxonomic review of the genusXylotrechus(Coleoptera:Cerambycidae:Cerambycinae)in Korea with a newly recorded species [J].Kor J Appl Entomol,2010,49(2):69-82.
[6] 丁玉洲,王业忠,吴六一,等.国内新发生的灾害性经济林害虫[J].安徽农业大学学报,2000,27(1):29-32.
[7] Chen J M,Qiao H L,Chen J,et al.Observation of antennal sensilla inXylotrechusgrayii(Coleoptera:Cerambycidae)with scanning electron microscopy [J].Microsc Res Tech,2014,77(4):264-273.
[8] 刘清琪,张俊林,李庆水,等.咖啡虎天牛为害金银花的初步研究[J].中药材,1981,4(2):18-19.
[9] 卫云,周曙明.山东金银花栽培技术[J].山东农业科学,1986,18(5):49-51.
[10] 余虹.金银花的病虫防治[J].四川农业科技,2003(5):26.
[11] 孙莹.金银花主要害虫发生和防治技术的研究[D].泰安:山东农业大学,2013.
[12] 王菊英.锈色粒肩天牛寄主选择机制初步研究[D].泰安:山东农业大学,2011.
[13] 诸葛飘飘.杨树云斑天牛成虫寄主定位中的信息化学物质[D].武汉:华中农业大学,2009.
[14] 郝德君,杨剑霞,戴华国.松墨天牛化学生态学[J].生态学杂志,2008,27(7):1227-1233.
[15] 王紫薇,徐华潮,张娓娓,等.光肩星天牛对寄主的选择及主要寄主挥发物的化学成分分析[J].浙江农林大学学报,2016,33(4):558-563.
[16] 李会平,黄大庄,王志刚,等.杨树形态特征、组织结构与光肩星天牛危害的关系[J].东北林业大学学报,2004,32(6):111-112.
[17] 刘博,徐华潮,孟俊国,等.视觉因素在松墨天牛定位寄主和交配中的作用[J].浙江农林大学学报,2012,29(4):617-620.
[18] 杨桦,王保新,杨伟,等.视觉在云斑天牛成虫选择寄主和寻找配偶中的作用[J].植物保护,2014,40(5):22-26.
[19] Yv F L,Hai X X,Wang Z G,et al.Integration of Visual and Olfactory Cues in Host Plant Identification by the Asian Longhorned Beetle,Anoplophoraglabripennis(Motschulsky)(Coleoptera:Cerambycidae)[J].PLoS One,2015,10(11):e0142752.
[20] Yang H,Wang B X,Yang W,et al.The Role of Vision in Host-Orientation and Mate-Finding Behaviors of the White-Striped Longhorned Beetle,Batoceralineolata[J].Entomol News,2017,126(5):358-371.
[21] 赵常胜.梣天牛生物学特性初步观察[J].中国森林病虫,2005,24(1):11-13.
[22] 缴丽莉,司永忠,董杰.石家庄地区桑天牛防治方法[J].现代农村科技,2014(19):25.
[23] 徐天,嵇保中,张琼岛,等.三清山高海拔栎林云斑白条天牛危害调查[J].中国森林病虫,2010,29(5):23-25.
[24] 陈德兰.杨树橙斑白条天牛的生物学特性[J].福建林学院学报,2010,30(2):137-140.
[25] 余黎红,陈国利,刘国军.双斑锦天牛的生物学特性及防治[J].植物保护,2007,33(2):108-110.
[26] 许畴,杨正德,贺永乾.梨眼天牛的生物学特性观察及其防治[J].西北林学院学报,2007,22(5):109-110.
[27] 刘东明,高泽正,邢福武.榕八星天牛生物学特性及其防治[J].中国森林病虫,2003,22(6):10-12.
[28] 张琼岛,嵇保中,徐天,等.栎旋木柄天牛取食与产卵选择性研究[J].应用昆虫学报,2011,48(3):626-633.
[29] 曹清,韦天雨,冯玉琴,等.园斑紫天牛生物学特性观察初报[J].河南林业科技,2009,29(3):47-48.
[30] 王跃,山钢,宋道军,等.花椒虎天牛的生物学特性及危害研究[J].四川林业科技,1999,20(3):38-40.
[31] 呼声久,孙丽娜,任炳生.桑虎天牛的初步研究[J].昆虫学报,1975,18(1):57-65.