杨-桤短期混交的林地土壤养分异质性
2018-05-11张贾宇徐叶宁陶慧颖鄂晓伟田野唐罗忠
张贾宇,徐叶宁,陶慧颖,鄂晓伟,田野,唐罗忠
南京林业大学林学院//南京林业大学南方现代林业协同创新中心,江苏 南京 210037
中国杨树(PopulusL.)人工林面积居世界第一(Isebrands et al.,2014)。为满足社会急迫的木材需求,杨树人工林多采取纯林连作的经营模式,出现了较为严重的地力衰退现象(王楚荣等,1999;张桥等,2004)。杨树生长迅速与其对土壤氮素(N)供应的高要求直接相关,因此,如何有效提高土壤氮素供应是维持杨树人工林长期立地生产力的关键。除施肥外,与固氮树种混交通常被认为是解决这一问题的一种有效而经济节约的方式。
目前就主要造林树种与固氮树种的混交方面已开展了一系列的研究,研究的侧重点主要在土壤养分状况改善、立地生产力提高以及养分在树种间的转移等方面(Kooch et al.,2016;Paula et al.,2015;Thurm et al.,2016)。固氮树种的引入可以提高林地土壤的N供应能力、加快主要树种凋落物分解、提高土壤微生物活性等(Mortimer et al.,2015;Teklehaimanot et al.,2007;Wu et al.,2014),从而提高主要树种的生产力(Binkley et al.,1992)。多数研究认为,固氮树种的养分改善效应通常需在混交较长时间后才能体现出来,如Mao et al.(2010)发现半干旱地区杨树与沙棘(Hippophae rhamnoides)混交5年后效果并不明显,直至混交15年后沙棘的促进作用才有所体现。但也有研究表明,混交的效应在较短时间内即可体现(Koupar et al.,2011;Sayyad et al.,2006)。此外,引入固氮树种在改善土壤氮素供应的同时,也有可能影响土壤氮、磷等养分供应的平衡,改变林地土壤的养分生态化学计量学特征(周正虎等,2016),从而影响林木的生长和林地生产力。比如,沈国舫等(1998)通过盆栽试验研究,认为沙地杨树与刺槐(Robinia pseudoacacia)混交提高林地生产力的原因一方面在于林地土壤N供应水平的改善,另一方面与刺槐和杨树间N、P养分的互补和转移也紧密关联。总体而言,对于固氮树种的引入能否在短期内对目的树种的氮素供应和生长产生影响目前并无定论,而针对短期混交对林地养分的生态化学计量的研究相对较少(周丽等,2014)。从林地微域角度看,固氮树种的引入有可能通过凋落物归还和根系作用在短期内改变林地局部的养分含量和平衡状况,并通过目的树种根系扩张改善其养分供应。
江南桤木(Alnus trabeculosa)是中国长江中下游地区广泛分布的一种乡土固氮树种,具有良好的固氮能力,喜湿速生,材性优良(邓廷秀,1984;王军辉等,2001),是中纬度平原地区营造杨树混交林的理想伴生树种。本论文以具有不同混交比例的杨树和江南桤木混交林为研究对象,对短期内混交林固氮树种所产生的养分效应进行调查,探讨混交林在林分形成初期所产生的区域养分异质性和养分生态化学计量学特征,以期为杨树混交林的培育提供参考。
1 材料与方法
1.1 研究地概况
研究地位于江苏省宿迁市泗洪县陈圩林场(N33°32′,E118°36′),地处洪泽湖西岸,属中纬度暖温带半湿润气候区。年平均气温 14.4 ℃,无霜期197 d左右,全年日照时间为2250~2350 h,年平均降水量为972.5 mm。土壤母质为洪泽湖淤积土,土壤质地为中壤至轻粘,透气透水性较差。
于2012年3月在杨树人工纯林的采伐迹地上营造混交试验林。造林株行距为3 m × 3 m,混交树种为南林 95 杨(P.×euramericana‘Nanlin-95’)和江南桤木,均为1年生苗。林分按照3种混交比例进行行状混交,分别为杨树-桤木2∶1混交(M1)、杨树-桤木1∶1混交(M2)、杨树-桤木1∶2混交(M3),并以杨树纯林(CK)作为对照。林分营造后未进行任何其他管理。
1.2 试验设计和样品采集
2016年6月下旬,在每种混交林分中设置3个重复小区,每个小区由3行3列共9株树围成,面积为36 m2。杨树纯林(CK)和杨树-桤木1∶1混交林分(M2)中树种均匀分布,不存在与固氮树种相对位置的差异,因此在每个重复小区内分别随机选取1个采样点进行取样。杨树-桤木2∶1混交林分(M1)和杨树-桤木 1∶2混交林分(M3)中由于存在杨树和桤木的非均匀分布,为了探讨杨树和桤木相对位置的不一致所产生的影响,在采样时划分成杨树与杨树、杨树与桤木、桤木与桤木区域,分别设置了不同的样点并分开取样。其中,M1中杨树与杨树行间围成区域所取的土壤样品记为M11,杨树与桤木行间所取的土壤样品记为M12;M3中杨树与桤木行间围成区域所取的土壤样品记为M31,桤木与桤木行间土壤样品记为M32。所有取样点均分布于树冠投影的边缘区域,以排除根际效应可能带来的干扰。每个取样点按土层深度采集0~10 cm和10~20 cm的土壤样品。采集的土壤样品去除明显的粗大杂质后放入冰盒暂存,并及时带回实验室。新鲜样品过2 mm筛,去除石砾、根系等杂物,并混合均匀。取一部分置于4 ℃冰箱中保存,用于 pH、铵态氮和硝态氮含量及有效磷测定。另取一部分风干土壤磨细过0.1 mm筛,用于土壤有机碳、全氮和全磷含量测定。
1.3 测定方法
土壤pH值采用电位计法测定(水土比2.5∶1)。土壤有机碳含量采用水合热重铬酸钾氧化-比色法(鲁如坤,2000)测定。土壤全氮含量采用浓硫酸消煮-靛酚蓝比色法测定。土壤全磷含量采用浓硫酸和高氯酸消煮-钼锑抗比色法测定。土壤硝态氮含量采用2 mol·L-1KCL浸提-酚二磺酸比色法测定。土壤铵态氮含量采用2 mol·L-1KCL浸提-靛酚蓝比色法测定。土壤有效磷含量采用0.5 mol·L-1碳酸氢钠(pH 8.5)浸提-钼锑抗比色法测定。土壤全氮、无机氮、全磷及有效磷含量的具体测定参照美国土壤学会的土壤分析测定方法(Sparks et al.,1996)。
1.4 数据处理
采用Microsoft Office 2013对试验各项指标数据进行计算处理;采用SPSS 21.0对各指标数据进行单因素方差分析(One-way ANOVA),采用最小显著差数法(least significant difference,LSD)进行多重比较;采用SigmaPlot 12.5进行图形绘制。
2 结果与分析
2.1 林地土壤pH值的变化
4种林分0~10 cm和10~20 cm土层土壤pH值的变化范围分别为5.76~7.65和6.24~7.29,两土层均以杨树-桤木 2∶1混交林分的杨树树行之间(M11)土壤 pH值最高,以杨树纯林(CK)林地土壤最低(图1)。引入桤木进行混交后0~10 cm土壤的 pH值分别显著升高了 32.66%、23.41%、24.10%、11.62%和9.16%,杨树-桤木2∶1混交(M1)和杨树-桤木1∶1混交(M2)林分的10~20 cm土壤pH值也均显著高于杨树纯林(CK)。由此可见,引入桤木混交5年一定程度上提高了林地表层土壤的pH值,但是,其提高效应随着桤木混交比例的增加而有所下降。由图1还可看出,杨树-桤木2∶1混交林分(M1)中0~10 cm土层的pH值出现了明显的异质性,两行杨树间(M11)的土壤 pH值比杨树与桤木行围成区域(M12)土壤 pH值显著高出7.49%。
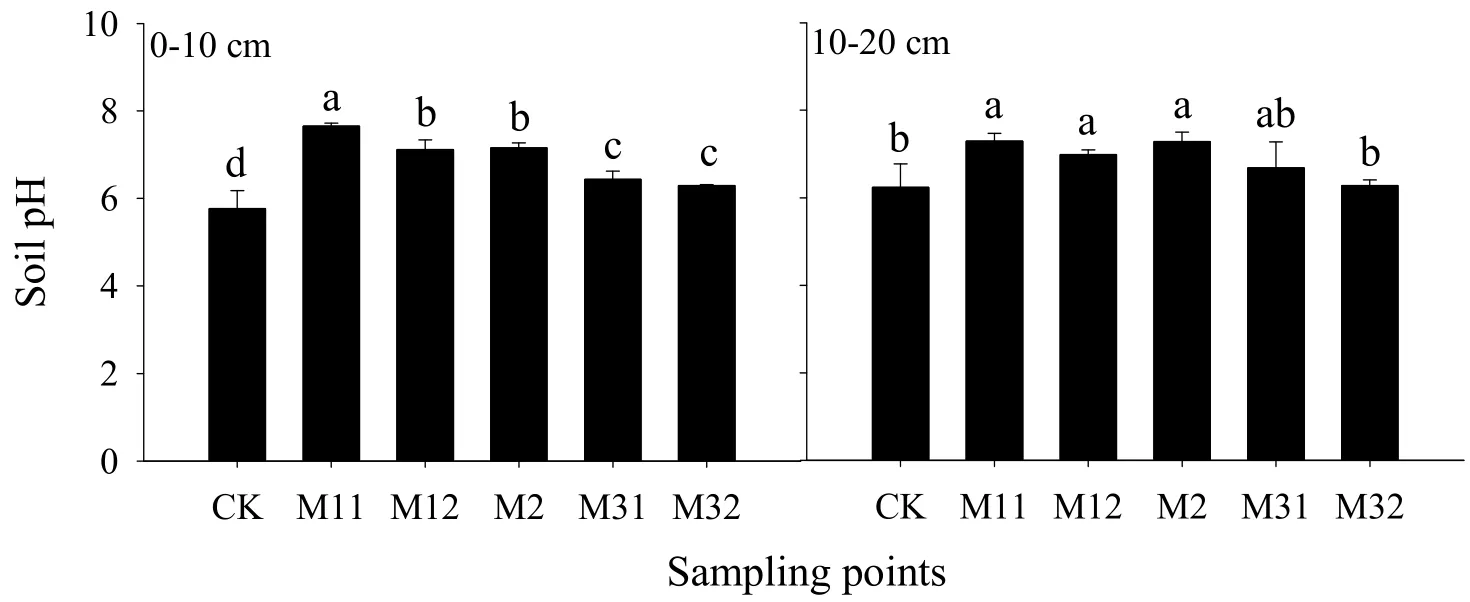
图1 不同混交比例林分中与杨树和桤木相对不同位置的土壤pH值Fig.1 Soil pH of different locations in the plantations with different mixing ratio of poplar and alder
2.2 林地土壤有机碳、全氮及全磷含量

图2 不同混交比例林分中与杨树和桤木相对不同位置的土壤有机碳、全氮及全磷含量Fig.2 Soil organic carbon, total nitrogen, and total phosphorus contents of different locations in the plantations with different mixing ratio of poplar and alder
由图2可知,4种林分0~10 cm土层土壤的有机碳、全氮及全磷含量整体上略高于10~20 cm土层。与杨树纯林(CK)相比,引入桤木进行混交并未显著提高林地土壤有机碳和全氮含量,但全磷含量有升高趋势。土壤有机碳含量均在15 g·kg-1左右波动;而与有机碳相比,混交林分的土壤全氮和全磷含量在不同林分以及林分的不同位置间的变动幅度相对较大。0~10 cm和10~20 cm土层土壤全氮含量均以杨树-桤木2∶1混交林分中杨树与桤木围成的区域(M12)相对较高,分别为1.57 g·kg-1和1.29 g·kg-1,但与其他区域的差异性并不显著;而该区域0~10 cm土层土壤的全磷含量(0.50 g·kg-1)比杨树纯林(CK)显著增加了 31.47%。由此可见,引入桤木进行混交5年内未对土壤有机碳和全氮总量产生显著影响,但就土壤全氮和全磷而言,短期的混交已经导致了林地的不同微域产生了一定的变化,随着混交时间的延长,林内的养分异质性可能会更加明显。
2.3 林地土壤无机氮和有效磷含量变化
由图3可知,本研究中各林分土壤的无机氮均以硝态氮为主,占总无机氮含量的98%以上,表明试验地土壤可能存在强烈的硝化作用。铵态氮和硝态氮含量在不同混交比例林分及林分内不同区域均有一定的变化,尤其在0~10 cm土层差异更显著。林地土壤铵态氮含量随着桤木混交比例的增大有所上升,其中0~10 cm和10~20 cm土层以杨树-桤木1∶2混交林分中两行桤木围成的区域(M32)最大,分别为 0.60 mg·kg-1和 0.42 mg·kg-1,较纯林土壤分别显著增加了112.67%和80.87%。0~10 cm土层的土壤硝态氮含量以混交林略高于杨树纯林(CK),且杨树-桤木2∶1混交林分两排杨树围成区域(M11)土壤硝态氮含量较杨树纯林(CK)增加了19.70%,10~20 cm土层的硝态氮含量在不同混交比例林分及林分内不同区域间均没有表现出显著差异,在34.43~37.66 mg·kg-1之间波动。
林地 0~10 cm土层土壤有效磷含量以杨树-桤木1∶1混交林分(M2)最高,达6.27 mg·kg-1,分别较杨树-桤木2∶1混交林分由两排杨树围成的区域(M11)和杨树-桤木1∶2林分两排桤木围成区域(M32)显著高出57.14%和34.93%。10~20 cm土层土壤有效磷含量略低于 0~10 cm土层,杨树-桤木1∶2林分桤木与桤木围成区域(M32)较杨树-桤木2∶1林分两排杨树围成区域(M11)和杨树-桤木 1∶1林分(M2)分别显著高出 40.07%和34.64%。
2.4 林地土壤C∶N∶P生态化学计量

图3 不同混交比例林分中与杨树和桤木相对不同位置的土壤无机氮及有效磷含量Fig.3 Soil inorganic nitrogen and available phosphorus in four mixed poplar-alder plantations relative positio n

表1 不同混交比例林分中与杨树和桤木相对不同位置的土壤养分生态化学计量Table1 Ecological stoichiometry of soil organic C, N and P of different locations in the plantations with different mixing ratio of poplar and alder
不同混交比例林分及其林地不同区域土壤的C∶N∶P生态化学计量学特征见表1。由表1可知,试验林地0~10 cm土层土壤C∶N比和C∶P比整体上低于10~20 cm土层土壤,而N∶P比则略高于10~20 cm土层土壤。方差分析结果表明,0~10 cm土层土壤C∶N比表现为杨树-桤木2∶1混交林分的杨树和桤木行间(M12)显著低于杨树纯林(CK),但10~20 cm土层的土壤C∶N比在各林分及不同位置间均无显著差异,说明杨树与桤木混交后短期内对土壤C∶N比的影响主要体现在0~10 cm的表层土壤。杨树-桤木2∶1混交林分的杨树和桤木行间(M12)0~10 cm土层的土壤C∶P比除显著低于杨树纯林(CK)外,还显著低于杨树-桤木1∶2混交林分的杨树和桤木行间土壤(M31),而对于10~20 cm土层,杨树-桤木2∶1混交林分(M1)和杨树-桤木1∶1混交林分(M2)的土壤C∶P比均显著低于杨树纯林(CK)。与土壤C∶N比和C∶P比不同,各林分及不同位置的土壤N∶P在0~10 cm土层均无显著差异,而10~20 cm土层则表现为杨树-桤木 2∶1混交林分的杨树和桤木行间(M12)显著高于杨树-桤木1∶1混交林分(M2)。
3 讨论
本研究结果表明,引入桤木混交造林5年后显著提高了杨树人工林林地表层土壤的pH值(图1),这一结果与前人针对林木混交的研究结果基本一致(刘姝媛等,2013;宋蒙亚等,2014;赵燕波等,2015)。杨树人工林引入桤木进行混交后,凋落物由原来单一的杨树落叶变为杨树与桤木的混合凋落物,林地分解群体的底物多样性提高,分解速度显著加快(Gartner et al.,2004;林开敏等,2001);此外,与杨树纯林(CK)相比,由于混交林树种间的养分转移(贾黎明等,1997),桤木根瘤固定的氮部分转移给杨树后,也可以提高杨树凋落物的养分质量,导致凋落物的分解进一步加快(陈国平等,2014),从而加速凋落物盐基离子的释放和归还(严海元等,2010),进而有效提高土壤pH值。本研究还发现,随着桤木混交比例的增加,其提升林地土壤pH值的效应有所下降,在杨树-桤木2∶1混交林分中(M1),由两排杨树围成的区域0~10 cm土层的土壤pH值甚至显著高于由杨树和桤木围成的区域,这一现象可能与林地整体凋落物量的变化有关。与杨树相比,桤木的生物量较小,凋落物量也明显较少,桤木混交比例的增大可能会减少林地凋落物的归还总量,从而在一定程度上减少盐基离子的归还量,使土壤pH值升幅变小。土壤pH值是影响土壤微生物的组成和活性、有机物分解和养分转化的重要因子(Tian et al.,2008;Tian et al.,2013),由于混交而导致的林地土壤pH值的变化所产生的效应目前尚缺乏相应的研究,将来需在针对混交林的研究中加以重视。
引入桤木进行混交在短期内对土壤有机碳和全氮含量影响较小(图 2),这一结果与前人的研究结果也基本一致(Chen et al.,2016;Rothe et al.,2002)。土壤有机碳和全氮的含量均以0~10 cm土层高于10~20 cm土层,这一点与凋落物归还时主要集中在土壤表面有关。凋落物需要经过长时间的分解后,其释放的养分才能逐渐向更深的土壤层次移动(朱秋莲等,2013)。杨树-桤木混交林林地土壤的硝态氮含量较杨树纯林(CK)有升高趋势,而且在桤木混交比例越高的林分,在靠近桤木一侧,土壤铵态氮含量也有升高趋势,这是桤木固氮作用的体现,表明不同比例桤木在短期混交后即已导致林地微域出现养分的异质性。然而,由图3可知,硝态氮和铵态氮在不同混交比例林分的不同微域中的表现不一致,其原因可能与桤木的固氮作用以及铵态氮和硝态氮在土壤中的行为方式有关。固氮树种所固定的氮在生长期主要以铵态氮的形式释放(何兴元等,1999),带正电荷的铵态氮容易被土壤胶体吸附,从而不容易移动,因此土壤铵态氮的含量以杨树-桤木1∶2混交林分中由两行桤木围成的区域(M32)最高(图 3)。本试验林地偏中性的土壤pH值还有利于硝化作用和硝态氮的生成(蔡祖聪等,2009),固氮作用所产生的大量铵态氮很容易转化为硝态氮;由于硝态氮在土壤中很容易随水流动,生成的硝态氮在杨树强烈的蒸腾吸水作用下很容易随着土壤液流转移到以杨树占优势的区域,导致杨树-桤木2∶1混交林分由两排杨树围成区域(M11)中土壤硝态氮含量最高。
从土壤 P的研究结果可知,杨树-桤木混交林0~10 cm土层土壤的全磷含量较杨树纯林(CK)有升高趋势,且以杨树-桤木2∶1混交林分中杨树与桤木围成的区域(M12)含量略高(图 2),而土壤有效磷含量则以杨树-桤木1∶1混交林分(M2)的林地0~10 cm土层土壤略高,说明混交对土壤磷含量及其供应也有一定的影响。这一结果可能也与混交引起的凋落物的分解和养分归还速率的加快有关。如陈琴(2012)通过室内控制条件下杨树和桤木混合凋落物的分解研究得出,虽然桤木凋落物中磷含量低于杨树,但两种凋落物混合分解时的分解速率和后期磷的释放速率明显高于单独杨树或桤木落叶。贾黎明等(1998)在研究沙棘与杨树凋落物混合分解时也发现了类似规律,混合凋落物分解速度更快,并且导致靠近桤木一侧土壤全磷含量更高。Hu et al.(2006)对杉木(Cunninghamia lanceolata)与其他树种的凋落物进行的为期2年的混合分解研究结果也表明,杉木与桤木凋落物混合后会增加微土壤生物活性,从而加快土壤磷的生物转化和有效磷的释放。此外,贺伟等(2003)通过杨树和刺槐的根箱混交试验发现,杨树可通过根系接触或根系分泌物将磷素转移到刺槐体内,这或许也是本试验靠近桤木侧土壤有效磷较高的可能原因之一。
土壤 C∶N∶P生态化学计量学特征是评价林地土壤在养分供应的平衡性方面的重要指标。本研究中,各林分土壤 C∶N比的变化范围基本接近Post et al.(1985)对全球土壤及 Tian et al.(2010)对中国土壤 C∶N比的报道,土壤 N∶P比也与Cleveland et al.(2007)所汇总的全球不同区域森林土壤的结果相近,但土壤C∶P比和N∶P比则整体上低于Tian et al.(2010)所报道的全国土壤平均值,这一结果可能主要与土壤P的含量有关。土壤中的P主要来自于成土母质的风化、有机物的归还以及施肥,受气候、土壤的熟化程度、植被的演替阶段以及土地的利用历史的显著影响。本试验的混交林在杨树采伐迹地上营建,上一代的杨树纯林在幼龄期曾进行数年的林农复合经营,有一定的肥料施用历史。由于磷在土壤中移动性差,不容易流失,因此所施用的肥料中的磷素可能在林地土壤中有一定的积累,这是本试验林土壤C∶P比和N∶P比低于全国土壤平均值的可能原因之一。引入桤木混交5年后,混交林分0~10 cm土层土壤C∶N比和 C∶P比较纯林有明显的下降趋势,与黄宇等(2004)所研究的杉木与桤木混交的效果相一致。固氮树种桤木的引入在短期内改变了混交林分局部区域表层土壤的C、N、P平衡,其中杨树-桤木2∶1混交林分的杨树与桤木行间(M12)的土壤C∶N比和C∶P比显著低于纯林(CK),说明短期混交可以提高局部土壤的 N、P供应能力(Giardina et al.,1995;李天平,2015),这可能是因为固氮树种的固氮作用和混合凋落物的分解加快,从而释放出更多的N、P养分(何兴元等,1999)导致。本研究中各林分及林内不同区域0~10 cm土层土壤的N∶P比并不存在显著差异,但均明显低于Cleveland et al.(2007)所认为可能存在磷限制的土壤N∶P比(5.89),因此,与土壤N的供应水平相比,本研究林地在一定程度上可能更容易受到土壤磷素的限制。磷在土壤中的动态和有效性与土壤pH值和根系行为直接相关,受土壤P的转化特性显著影响,在将来的研究中也需进一步加以关注。
总体上,本研究结果表明,引入桤木与杨树混交后,在短期内即可以导致不同混交比例林分内不同区域的pH值、养分元素含量及生态化学计量学特征产生一定的异质性,在一定程度上改变了林地的养分供应和养分平衡。本课题组前期的研究还发现,在杨-桤混交林内,杨树可以通过积极扩张根系,扩大吸收根分布范围的策略来提高养分利用效率,从而获取更多养分(章晴,2016)。因此,由于引入桤木进行混交引起的林地土壤养分异质性,特别是土壤养分提高的效果,虽然发生在林分内的不同区域,但是可以通过杨树根系的扩张来加以利用,从而在短期内即对杨树的生长产生促进效果。当然,相应的效果还需要通过更为精细化的生长监测以及杨树的生理特征的改变研究进一步加以验证和评价,并在以后的研究中补充有关凋落物归还量的研究。
4 结论
混交5年后,各混交比例杨-桤混交林0~10 cm土壤pH值均显著高于杨树纯林,但随桤木混交比例的增大,其提高土壤pH值的效应减弱,且0~10 cm土壤pH值在杨-桤2∶1混交林分中出现明显的异质性。短期混交对杨树林土壤有机碳和全氮影响较小,但全磷含量略有提升。试验林地土壤无机氮以硝态氮为主,各混交比例林分间不存在显著差异,但在同一混交林分中铵态氮含量有一定的异质性,靠近桤木行的区域其土壤铵态氮含量整体上高于靠近杨树行的区域,且在杨树-桤木1∶2混交林的10~20 cm土壤中表现显著。短期混交对土壤C∶N比的影响主要体现在0~10 cm的表层土壤;混交林分0~10 cm土层土壤C∶N比和C∶P比较纯林有明显下降趋势,表明短期混交后一定程度上提高了林地局部土壤的N、P供应能力。
参考文献:
BINKLEY D, SOLLINS P, BELL R, et al. 1992. Biogeochemistry of adjacent conifer and alder-conifer stands [J]. Ecology, 73(6):2022-2033.
CHEN Y Q, YU S Q, LIU S P, et al. 2016. Reforestation makes a minor contribution to soil carbon accumulation in the short term: evidence from four subtropical plantations [J]. Forest Ecology and Management,384: 400-405.
CLEVELAND C C, LIPTZIN D. 2007. C∶N∶P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? [J].Biogeochemistry, 85(3): 235-252.
GARTNER T B, CARDON Z G. 2004. Decomposition dynamics in mixed-species leaf litter [J]. OIKOS, 104(2): 230-246.
GIARDINA C P, HUFFMAN S, BINKLEY D, et al. 1995. Alders increase soil phosphorus availability in a Douglas-fir plantation [J]. Canadian Journal of Forest Research, 25: 1652-1657.
HU Y, WANG S, ZENG D. 2006. Effects of single Chinese fir and mixed leaf litters on soil chemical, microbial properties and soil enzyme activities [J]. Plant and Soil, 282(1): 379-386.
ISEBRANDS J G, RICHARDSON J. 2014. Poplars and willows - trees for society and the environment [M]. Rome: FAO and CABI: 634.
KOOCH Y, ROSTAYEE F, HOSSEINI S M. 2016. Effects of tree species on topsoil properties and nitrogen cycling in natural forest and tree plantations of northern Iran [J]. Catena, 144: 65-73.
KOUPAR S A M, HOSSEINI S M, MODIRRAHMATI A, et al. 2011.Effect of pure and mixed plantations ofPopulus deltoideswithAlnus subcordataon growth, nutrition and soil properties [J]. Asian Journal of Chemistry, 23(3): 950-954.
MAO R, ZENG D H, AI G Y, et al. 2010. Soil microbiological and chemical effects of a nitrogen-fixing shrub in poplar plantations in semi-arid region of Northeast China [J]. European Journal of Soil Biology, 46(5): 325-329.
MORTIMER P E, GUI H, XU J, et al. 2015. Alder trees enhance crop productivity and soil microbial biomass in tea plantations [J]. Applied Soil Ecology, 96: 25-32.
PAULA R R, BOUILLET J P, TRIVELIN P C O, et al. 2015. Evidence of short-term belowground transfer of nitrogen fromAcacia mangiumtoEucalyptus grandistrees in a tropical planted forest [J]. Soil Biology and Biochemistry, 91: 99-108.
POST W M, PASTOR J, ZINKE P J, et al. 1985. Global patterns of soil nitrogen storage [J]. Nature, 317(6038): 613-616.
ROTHE A, CROMACK K, RESH S C, et al. 2002. Soil carbon and nitrogen changes under Douglas-fir with and without red alder [J]. Soil Science Society of America Journal, 66(6): 1988-1995.
SAYYAD E, HOSSEINI S M, MOKHTARI J, et al. 2006. Comparison of growth, nutrition and soil properties of pure and mixed stands ofPopulus deltoidesandAlnus subcordata[J]. Silva Fennica, 40(1):27-35.
SPARKS D L, PAGE A L, HELMKE P A, et al. 1996. Methods of soil analysis. Part 3-chemical methods [M]. Madison: Soil Science Society of America: 869-1184.
TEKLEHAIMANOT Z, MMOLOTSI R M. 2007. Contribution of red alder to soil nitrogen input in a silvopastoral system [J]. Biology and Fertility of Soils, 43(6): 843-848.
THURM E A, PRETZSCH H. 2016. Improved productivity and modified tree morphology of mixedversuspure stands of European beech(Fagus sylvatica) and Douglas-fir (Pseudotsuga menziesii) with increasing precipitation and age [J]. Annals of Forest Science, 73(4):1047-1061.
TIAN H Q, CHEN G S, ZHANG C, et al. 2010. Pattern and variation of C∶N∶P radios in China's soils: a synthesis of observational data [J].Biogeochemistry, 98(1-3): 139-151.
TIAN Y, HAIBARA K, TODA H, et al. 2008. Microbial biomass and activity along a natural pH gradient in forest soils in a karst region of the upper Yangtze River, China [J]. Journal of Forest Research, 13(4):205-214.
TIAN Y, TAKANASHI K, TODA H, et al. 2013. pH and substrate regulation of nitrogen and carbon dynamics in forest soils in a karst region of the upper Yangtze River basin, China [J]. Journal of Forest Research, 18(3): 228-237.
WU F Z, PENG C H, YANG W Q, et al. 2014. Admixture of alder (Alnus formosana) litter can improve the decomposition of eucalyptus(Eucalyptus grandis) litter [J]. Soil Biology and Biochemistry, 73(6):115–121.
蔡祖聪, 赵维. 2009. 土地利用方式对湿润亚热带土壤硝化作用的影响[J]. 土壤学报, 46(5): 795-801.
陈国平, 程珊珊, 丛明旸, 等. 2014. 3种阔叶林凋落物对下层土壤养分的影响[J]. 生态学杂志, 33(4): 874-879.
陈琴. 2012. 杨树-桤木落叶混合分解的养分释放动态及其对土壤养分特性的影响[D]. 南京: 南京林业大学: 18-19.
邓廷秀. 1984. 水土保持固氮树种-桤木、刺槐、山合欢、马桑、紫穗槐[J]. 中国水土保持, (8): 22-24.
何兴元, 赵淑清, 杨思河, 等. 1999. 固氮树种在混交林中的作用研究Ⅲ. 固氮树种凋落物分解及 N的释放[J]. 应用生态学报, 10(4):404-406.
贺伟, 贾黎明, 赫宝刚, 等. 2003. 混栽杨树-刺槐间磷素养分转移途径的研究[J]. 应用生态学报, 14(4): 481-486.
黄宇, 冯宗炜, 汪思龙, 等. 2004. 杉木与固氮和非固氮树种混交对林地土壤质量和土壤水化学的影响[J]. 生态学报, 24(10): 2192-2199.
贾黎明, 方陆明. 1998. 杨树刺槐混交林及纯林枯落叶分解[J]. 应用生态学报, 9(5): 463-467.
贾黎明, 翟明普, 胡延杰, 等. 1997. 杨树刺槐种间氮素养分转移[C]//沈国舫, 翟明普. 混交林研究. 北京: 中国林业出版社: 42-49.
李天平. 2015. 湘北丘陵区混交阔叶林不同树种土壤C、N、P生态化学计量学特征研究[D]. 长沙: 中南林业科技大学: 25-28.
林开敏, 洪伟, 俞新妥, 等. 2001. 杉木与伴生植物凋落物混合分解的相互作用研究[J]. 应用生态学报, 12(3): 321-325.
刘姝媛, 胡浪云, 储双双, 等. 2013. 3种林木凋落物分解特征及其对赤红壤酸度及养分含量的影响[J]. 植物资源与环境学报, 22(3): 11-17.
鲁如坤. 2000. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社:109-110.
沈国舫, 贾黎明, 翟明普. 1998. 沙地杨树刺槐人工混交林的改良土壤功能及养分互补关系[J]. 林业科学, 34(5): 12-20.
宋蒙亚, 李忠佩, 刘明, 等. 2014. 不同林地凋落物组合对土壤速效养分和微生物群落功能多样性的影响[J]. 生态学杂志, 33(9): 2454-2461.
王楚荣, 陈红跃, 许炼烽. 1999. 混交林的改土效益及其在防止地力衰退上的应用[J]. 土壤与环境, 8(1): 58-60.
王军辉, 顾万春, 夏良放, 等. 2001. 桤木种源(群体)/家系材性性状的遗传变异[J]. 林业科学研究, 14(4): 362-368.
严海元, 辜夕容, 申鸿. 2010. 森林凋落物的微生物分解[J]. 生态学杂志, 29(9): 1827-1835.
张桥, 蔡婵凤. 2004. 森林土壤退化及其防治研究综述[J]. 生态环境,13(4): 677-680.
章晴. 2016. 杨-桤混交林根系碳分泌及其对根际土壤氮转化的影响[D].南京: 南京林业大学: 24-25.
赵燕波, 纪托未, 张丹桔, 等. 2015. 3个树种与巨桉混交土壤理化性质、凋落物量和养分含量特征[J]. 应用与环境生物学报, 21(5): 948-953.
周丽, 张卫强, 唐洪辉, 等. 2014. 南亚热带中幼龄针阔混交林生态化学计量特征[J]. 生态环境学报, 23(11): 1732-1738.
周正虎, 王传宽. 2016. 微生物对分解底物碳氮磷化学计量的响应和调节机制[J]. 植物生态学报, 40(6): 620-630.
朱秋莲, 邢肖毅, 张宏, 等. 2013. 黄土丘陵沟壑区不同植被区土壤生态化学计量特征[J]. 生态学报, 33(15): 4674-4682.
